

8.2. Факторы клеточной дифференциации
Основными факторами клеточной дифференцировки являются: 1) пролиферация клеток; 2) морфогенетические движения клеток и их пластов (амебоидное движение, иммиграция, инвагинация); 3) избирательная сортировка и адгезивность клеток; 4) гибель клеток; 5) избирательная активация генов и синтез типоспецифических белков (актин, миозин, гемоглобин) в исходно однородных клетках. Морфологически выявляемой дифференцировке (формообразованию) предшествует биохимическая дифференцировка, т.е. изменение обмена веществ.
Пролиферация клеток (размножение клеток) лежит в основе развития всех органов. Благодаря ей образуется определённое количество клеток, создающее возможность для возникновения различий между клетка-

Рис.
116.
Морфогенетические
движения клеток при гаструляции
и
первичном органогенезе у ланцетника
ми группы, т.е. их дифференциации. Делящиеся клетки могут располагаться без видимого порядка или концентрироваться в особых матричных зонах зачатков органов.
Морфогенетические движения клеток и их пластов представляют собой преимущественно активные перемещения клеток посредством механизма амебоидного движения. Траектория перемещения определяется чаще всего рельефом поверхности, по которой перемещается клетка (контактная ориентировка). Значительно реже встречается перемещение клеток по градиенту концентрации химических веществ (хемотаксис). Активные перемещения клеток способствуют доставке клеточного материала в соответствующую область зародыша (рис. 116). Основным отличием клеток разных зародышевых листков являются их различные морфогенетические движения, характер которых специфичен у разных групп животных. Эктодерма в ходе гаструляции, как правило, распластывается и, оставаясь на поверхности, окружает весь зародыш. Энтодерма, напротив, свёртывается в трубку - первичную кишку (рис. 116). У зародышей морского ежа она сворачивается внутрь полости бластулы. Мезодерма, образуясь на границе эк-
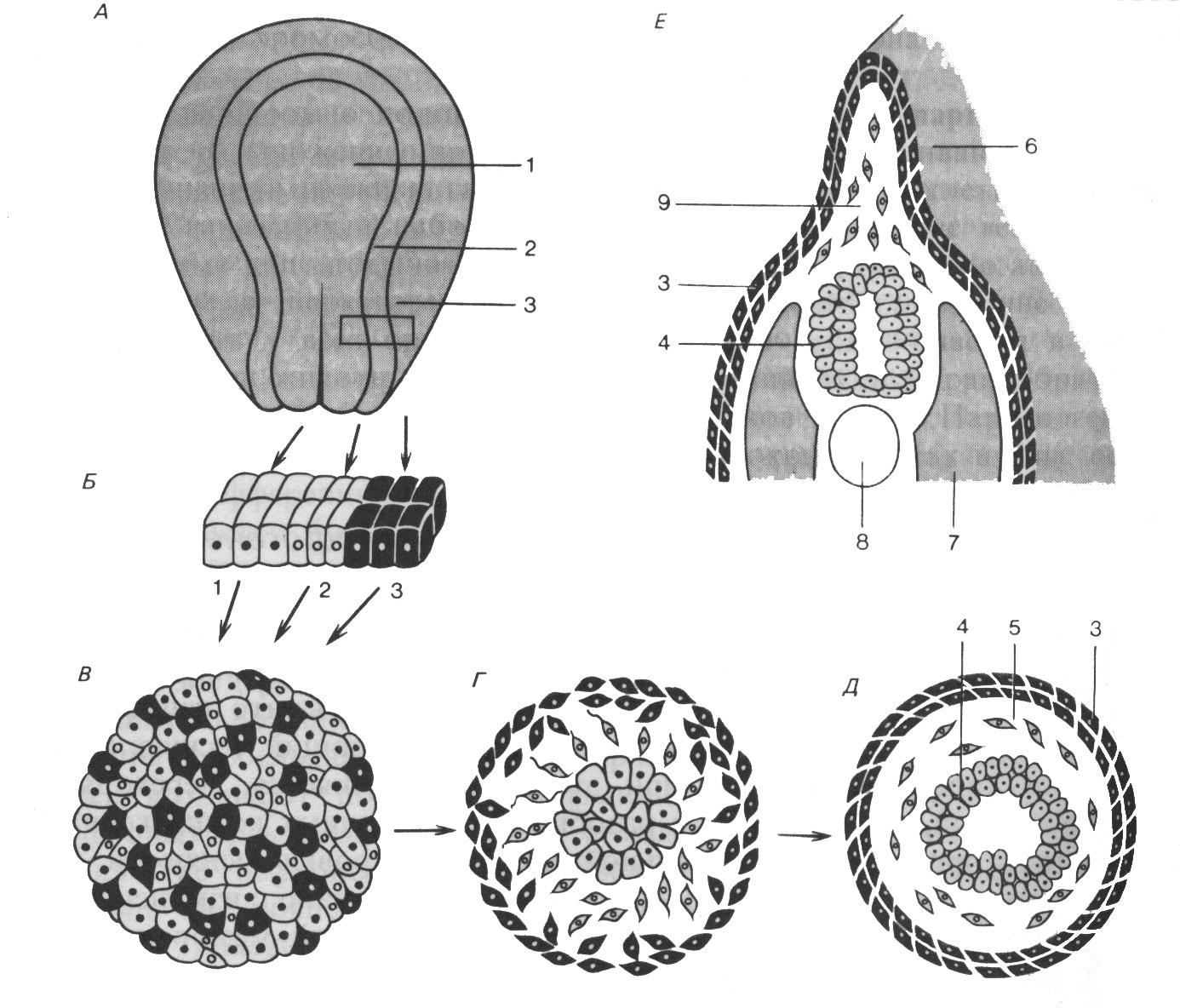
Рис.
117. Избирательная
сортировка и адгезивность клеток
как
механизм морфогенеза.
А – эксплантация
трёх типов клеток: 1 – нервная пластинка;
2 – нервный валик нейрулы амфибий; 3 –
эктодерма. Б – положение до диссоциации.
В – реагрегация в эксплантате.
Г –
начало дифференцировки. Д –
дифференцированная конечная стадия с
нервной трубкой (4), мезенхимой (5) и
эктодермой (3). Е – соответствующая
эксплантату (Д) область личинки: 6 –
край плавника; 7 – миотом; 8 – хорда; 9 –
мезенхима края плавника
тодермы и энтодермы, также вворачивается и проникает под эктодерму, отделяя её от энтодермы. На конечную судьбу клеток влияют контакты, возникающие по ходу их движения.
Избирательная сортировка и адгезивность заключается в выделении и объединении клеток одного зачатка из совокупности, содержащей клетки различных зачатков. Она свойственна клеточному материалу как зародышевых листков, так и отдельных органов (рис. 117). Объединение сходных клеток носит, по-видимому, вероятностный характер: межклеточные контакты образуются случайно, однако связи между однотипными клетками отличаются большей устойчивостью (более высокой адгезивностью). Более высокая адгезивность однотипных клеток и обеспечивает в конечном итоге формирование упорядоченно расположенных клеток одного зачатка (клеточного комплекса). В классических опытах Гельтфретера клетки были диссоциированы - отделены одна от другой, а затем перемешаны и вновь соединены (рис. 117). Через некоторое время, благодаря индивидуальным движениям клеток, клетки эктодермы оказались снаружи, энтодермы - внутри, а мезодермы - между ними. Описанное не свойственно только злокачественно перерождённым клеткам: они не выделяются из смеси с нормальными клетками, а потому могут прорастать в различные ткани и органы.
Многочисленными опытами показано, что наибольшей адгезивностью обладает мезодерма, наименьшей - эктодерма, а энтодерма занимает промежуточное между ними положение. Причём в действительности наибольшей взаимной адгезивностью обладают клетки эктодермы, но общая адгезивность эктодермы резко снижается из-за малой адгезивности наружной поверхности её клеток. То, что внутренняя поверхность эктодермы обладает наибольшей адгезивностью, способствует следующему: что мезодерма, более адгезивная, чем энтодерма, занимает промежуточное положение между эктодермой и энтодермой, контактируя с внутренней поверхностью эктодермы.
В связи с тем, что мезодерма менее адгезивна, чем внутренняя поверхность эктодермы, она «стремится» распространяться по эктодерме. Этим можно объяснить разрастание крыши первичной кишки и отчасти инвагинацию мезодермы. Позднее энтодерма будет распространяться по внутренней поверхности более адгезивной инвагинировавшей мезодермы, и, таким образом, будет достигнуто типичное расположение зародышевых листков. При удалении эктодермы мезодерма погружается в энтодерму. Следовательно, региональные различия в адгезивности способствуют возникновению сложной тканевой организации. Большое значение для развивающегося зародыша имеет неадгезивный наружный слой эктодермы. Он мешает зародышу «вывернуться наизнанку». Если его удалить, то энтодерма, имеющая теперь свободный край, может покрыть, разрастаясь, весь зародыш.
Развитие ряда органов предполагает гибель клеток, их локальных групп или части закладок органов. Такая гибель клеток запрограммирована, т.е. предопределена генетически. Гибель клеток часто происходит там, где в сплошных закладках вторично возникает полость, а также в тех случаях, когда стенки, разделяющие группы частично дифференцированных клеток, развиваются с образованием отверстия. Типичным примером такой гибели клеток является образование пальцев у позвоночных: на лопатовид-
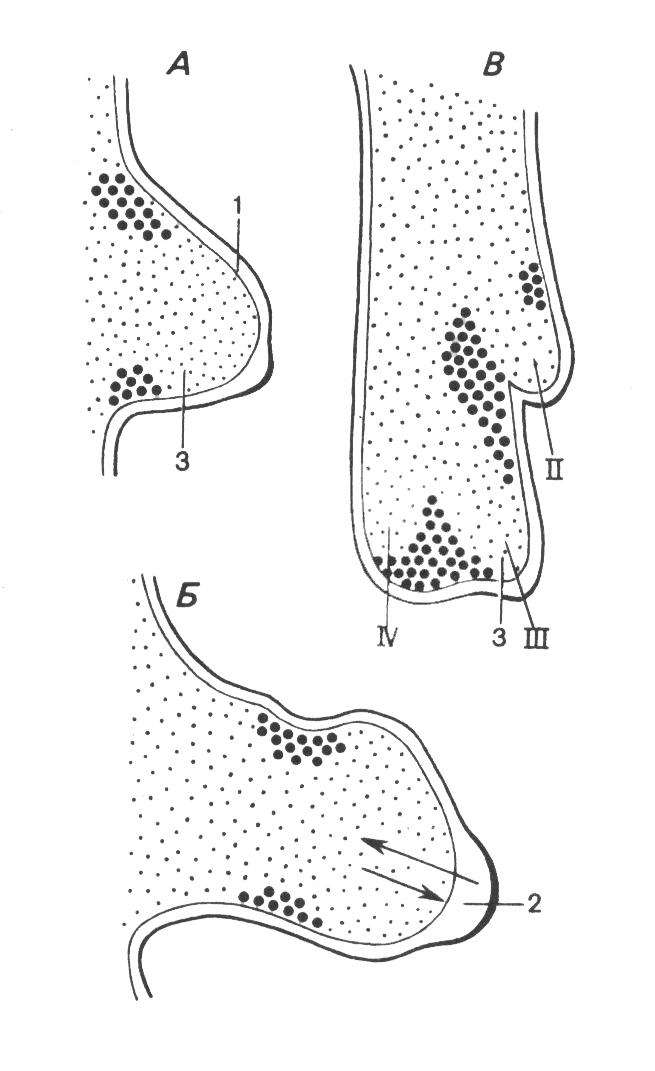
Рис. 118.
Запрограммированная
гибель клеток (области, помеченные
жирными точками) в закладке крыла
цыплёнка, а также взаимодействие
(стрелки) между эктодермой (1, белая) с
эпидермальным гребнем (2) и мезодермой
(3, мелкие точки). А – ранняя стадия. Б –
закладка в форме лопаточки с полностью
развитым эпидермальным гребнем. В -
подготовка к разделению лучей II,
III
и IV
дифференцирующихся фаланг
ном зачатке конечности возникают центры клеточной гибели, которые обеспечивают разделение лучей дифференцирующихся фаланг пальцев (рис. 118). При нарушении дифференцировки процессы, ведущие к запрограммированной гибели клеток, блокируются, в результате чего развиваются конечности со сросшимися пальцами или с перепонками между ними.
Формирование различий между однородными клетками начинается с возникновения между ними биохимических различий: в клетках начинают синтезироваться органические вещества различной природы, например, типоспецифические белки (миозин, опсин, гемоглобин), а также вещества углеводной и иной природы. Биохимические различия определяют в последующем функциональную специализацию клеток, особенности их морфологии: клетки, в которых синтезируется преимущественно миозин, дают начало мышечным волокнам; клетки, в которых накапливается опсин, становятся фоторецепторными клетками.
Все клетки зародыша имеют идентичный набор генов. Главной причиной возникновения между ними различий (дифференциации клеток) рассматривается избирательная (дифференциальная) активность генов: гены или их группы избирательно активируются (деблокируются) или, наоборот, инактивируются (блокируются).
В ходе дифференцировки
клеток зародыша наблюдается
последовательная смена активных
генов. Удобным объектом для наблюдения
этого явления служат гигантские
политенные хромосомы клеток некоторых
о рганов
насекомых, например, дрозофилы. Зоны
таких хромосом, содержащие активные
гены, на которых интенсивно синтезируется
РНК, выглядят как вздутия (пуфы).
Локализация пуфов по длине хромосомы
варьирует в зависимости от стадии
развития и тканевой принадлежности
клетки (рис. 119).
рганов
насекомых, например, дрозофилы. Зоны
таких хромосом, содержащие активные
гены, на которых интенсивно синтезируется
РНК, выглядят как вздутия (пуфы).
Локализация пуфов по длине хромосомы
варьирует в зависимости от стадии
развития и тканевой принадлежности
клетки (рис. 119).
П
Рис. 119. Участок
хромосомы с пуфом, где часть ДНК
раскручена для транскрипции, а остальная
ДНК туго свёрнута и недоступна для
РНК-синтезирующих
ферментов
При этом не происходят обязательно необратимые изменения клеточного ядра, что было продемонстрировано Дж. Гордоном: ядро дифференцированной клетки кишечника головастика изолировалось и вводилось в яйцеклетку лягушки, в которой путём ультрафиолетового облучения предварительно было убито собственное ядро; в ряде случаев такие яйца развивались во взрослых особей (рис. 120). Следовательно, вся информация, необходимая для нормального развития тотипотентной клетки, присутствует в ядре дифференцированной клетки, может вновь активироваться и использоваться для повторения процесса развития.