

10.2.2. Организация функционирования иммунной системы
Иммунная система имеет сложную организацию - для осуществления специфической функции задействовано множество различных клеточных популяций и растворимых факторов иммунитета. Клетки постоянно циркулируют в организме, погибают в процессе жизнедеятельности и воспроизводятся.
В зависимости от конкретной потребности специфическая функция иммунной системы может быть активирована либо подавлена (супрессирована). Однако любое реагирование иммунной системы осуществляется только при постоянном взаимодействии практически всех типов ее клеток, т.е. в условиях межклеточной кооперации. Раздражителем (активирующим сигналом) является антиген. В развитии любого иммунного реагирования прослеживается каскад последовательно сменяющихся этапов.
|
|
10.2.2.1. Взаимодействие клеток иммунной системы
Необходимым условием функционирования иммунной системы является тесная межклеточная кооперация,основу которой составляет рецептор-лигандное взаимодействие. Для связи между собой клетки используют различные дистантные растворимые факторы и прямой контакт.
Синтез растворимых факторов является одним из универсальных способов коммутации клеток между собой. К таковым относятся цитокины, которых в настоящее время известно более 25. Они представляют собой гетерогенное семейство разнообразных по структуре и функции биологически активных молекул, имеющих ряд общих свойств:
• как правило, цитокины не депонируются в клетке, а синтезируются после соответствующего стимула;
• для восприятия цитокинового сигнала клетка экспрессирует соответствующий рецептор, который может взаимодействовать с несколькими различными цитокинами;
• цитокины синтезируются клетками разных ростков, уровней и направлений дифференцировки;
• субпопуляции клеток иммунной системы различаются по спектру синтезируемых цитокинов и их рецепторов;
• цитокины обладают универсальностью, множественностью эффектов и синергизмом;
• цитокины могут воздействовать как на рядом расположенную клетку (паракринная регуляция), так и на сам продуцент (аутокринная регуляция);
• цитокиновая регуляция носит каскадный характер: активация клетки одним цитокином вызывает синтез другого;
• в подавляющем большинстве это короткодистантные медиаторы - их эффекты проявляются на месте выработки. Вместе с тем ряд провоспалительнъгх цитокинов (ИЛ-1, 6, α-ФНО и др.) могут оказывать системное действие.
Цитокины различаются по ведущей функциональной направленности:
• медиаторы доиммунного воспаления (ИЛ-1, 6,12, α-ФНОидр);
|
|
• медиаторы иммунного воспаления (ИЛ-5, 9, 10, γ-ИФН
и др.);
• стимуляторы пролиферации и дифференцировки лимфоцитов (ИЛ-2, 4, 13, трансформирующий фактор роста - β-ТФР
и др.);
• факторы роста клеток, или колониестимулирующие факторы
(ИЛ-3, 7, ГМ-КСФ и др.);
• хемокины, или клеточные хемоаттрактанты (ИЛ-8 и др.). Краткая характеристика некоторых цитокинов приведена в
табл. 10.3.
Прямое межклеточное взаимодействие основано на рецепции структур, экспрессированных на мембране клетки-оппонента. Для этого требуется достаточно продолжительный и стабильный контакт клеток. Такой способ коммутации используют Т-хелперы и Т-киллеры при анализе чужеродности презентированных структур. Механизм действия ко-стимулирующих факторов (пары CD40- CD40-лиганд, CD28-CD80, 86) также требует непосредственного контакта.
10.2.2.2. Активация иммунной системы
Активация иммунной системы подразумевает развитие продуктивной иммунной реакции в ответ на антигенное раздражение
Таблица 10.3. Характеристика основных цитокинов
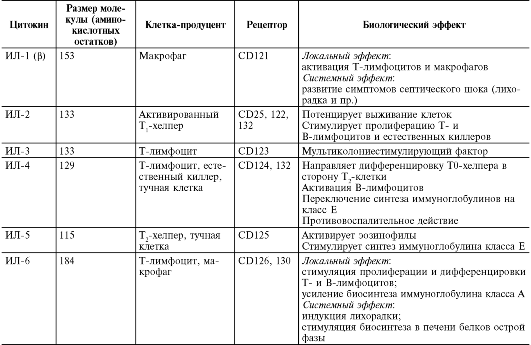 Продолжение
табл. 10.3
Продолжение
табл. 10.3
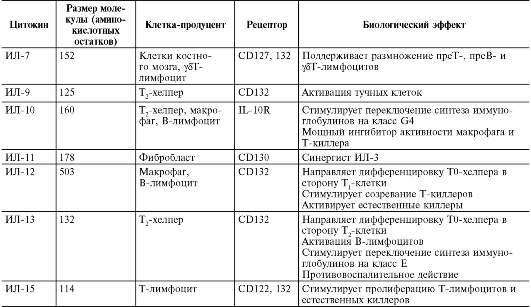 Продолжение
табл. 10.3
Продолжение
табл. 10.3
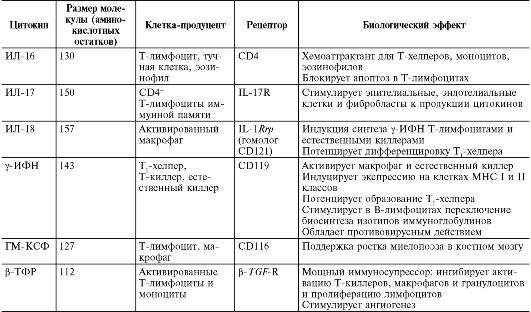 Окончание
табл. 10.3
Окончание
табл. 10.3
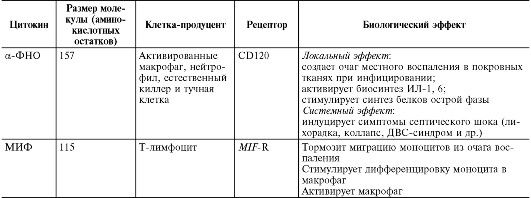 Примечание.
МИФ - миграцию ингибирующий фактор.
Примечание.
МИФ - миграцию ингибирующий фактор.
и появление продуктов деструкции тканей макроорганизма. Это сложный многоступенчатый процесс, требующий продолжительного времени для своей индукции - около 4 сут. Критическим событием является невозможность элиминации антигена факторами врожденного иммунитета в течение указанного срока.
Пусковым механизмом адаптивного иммунитета является распознавание «свой-чужой», которое осуществляют Т-лимфоциты при помощи своих прямых иммунорецепторов - TCR. В случае установления чужеродности биоорганической молекулы включается второй этап реагирования - запускается интенсивное тиражирование клона высокоспецифичных для антигена лимфоцитовэффекторов, способных прервать антигенную интервенцию. Это явление получило название «экспансия клона». Параллельно, но несколько позже пролиферации стимулируются дифференцировка иммунных лимфоцитов и формирование из него клеток иммунологической памяти, гарантирующих выживание в будущем.
|
|
Таким образом, продуктивная активация иммунной системы связана с размножением и дифференцировкой антигенореактивных клонов иммунокомпетентных клеток. Антигену в этом процессе отведена роль индуктора и фактора клональной селекции. Механизмы основных этапов активации иммунной системы рассмотрены ниже.
Активация Т-хелпера. Процесс (см. рис. 10.6) осуществляется при непосредственном участии АПК (дендритные клетки, В-лимфоциты и макрофаги). После эндоцитоза и процессинга антигена во внутриклеточных везикулах АПК встраивает образовавшийся олигопептид в молекулу MHC II класса и выставляет полученный комплекс на наружной мембране. На поверхности АПК также экспрессируются ко-стимулирующие факторы - молекулы CD40, 80, 86, мощным индуктором которых являются продукты разрушения покровных тканей на этапе доиммунного воспаления.
Т-хелпер при помощи молекул адгезии прочно соединяется с поверхностью АПК. Иммунорецептор Т-хелпера совместно с молекулой CD3 при поддержке ко-рецепторной молекулы CD4 взаимодействует с комплексом антиген-MHC II класса и анализирует чужеродность его структуры. Продуктивность рецепции зависит от ко-стимулирующих воздействий в парах CD28-CD80/86 и CD40- лиганд-CD40.
В случае признания чужеродности комплекса антиген-MHC II класса (точнее, «не своего») Т-хелпер активируется. Он экспресси-
рует рецептор к ИЛ-2 и начинает синтезировать ИЛ-2 и другие цитокины. Итогом активации Т-хелпера являются его размножение и дифференцировка в одного из своих потомков - T1- или Т2-хелпер (см. рис. 10.2). Любое изменение условий рецепции прекращает активацию Т-хелпера и может индуцировать в нем апоптоз.
Активация В-лимфоцита. Для активации В-лимфоцита (рис. 10.9) необходима суммация трех последовательных сигналов. Первый сигнал - результат взаимодействия молекулы антигена со специфичным для него BCR, второй - интерлейкиновый стимул активированного Т-хелпера и третий - результат взаимодействия ко-стимулирующих молекул CD40 с CD40-лигандом.
|
|
Активация инициирует размножение и дифференцировку специфичного для конкретного антигена В-лимфоцита (см. рис. 10.2). В итоге в пределах зародышевых (герминативных) центров лимфоидных фолликулов появляется клон специфических антителопродуцентов. Дифференцировка позволяет переключить биосинтез иммуноглобулинов с классов M и D на более экономные: G, A или Е (редко), повысить аффинность синтезируемых антител и образовать В-клетки иммунологической памяти или плазматические клетки.
Активация В-лимфоцита - весьма тонкий процесс. Отсутствие хотя бы одного из стимулов (нарушение межклеточной кооперации, неспецифичность рецептора В-лимфоцита или элиминация антигена) блокирует развитие антительного иммунного ответа.
Активация Т-киллера. Для исполнения надзорной функции Т-киллер вступает в тесный и прочный контакт с потенциальной
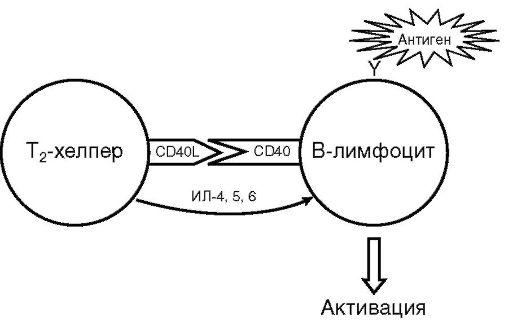 Рис.
10.9. Схема
активации В-лимфоцита (пояснения в
тексте)
Рис.
10.9. Схема
активации В-лимфоцита (пояснения в
тексте)
клеткой-мишенью, используя молекулы адгезии (см. рис. 10.8). Затем иммунорецептор Т-киллера (αβTCR)совместно с молекулой CD3 при поддержке ко-рецепторной молекулы CD8 взаимодействует с антигенным комплексом MHC I класса и анализирует его структуру. Обнаружение отклонений в пользу аллогенности активирует Т-киллер к экспрессии рецептора к ИЛ-2 и синтезу ИЛ-2 и высвобождение эффекторных молекул (перфорин, гранзимы, гранулизин) из цитоплазматических гранул в синаптическую щель межклеточного контакта.
Для адекватного развития клеточной формы иммунного ответа требуются активизирующие стимулы со стороны Т1-хелпера. Т-киллер может функционировать автономно, самостоятельно инициируя и поддерживая клонообразование за счет аутокринной стимуляции ИЛ-2. Однако это свойство реализуется редко.
10.2.2.3. Супрессия иммунного ответа
Супрессия или подавление иммунного ответа является физиологической реакцией организма, которая в норме завершает иммунный ответ и направлена на торможение экспансии антигенспецифических клонов лимфоцитов. В отличие от иммунологической толерантности, супрессии подвергается уже инициированное иммунное реагирование. Различают три механизма иммуносупрессии: уничтожение клонов иммунокомпетентных клеток, торможение активности иммунокомпетентных клеток, элиминация антигенного стимула.
|
|
Устранить иммунокомпетентные клетки можно путем апоптоза. При этом элиминации подвергаются следующие группы клеток:
• терминально дифференцированные лимфоциты, завершившие свою биологическую программу;
• активированные лимфоциты, не получившие антигенного стимула;
• «изношенные» лимфоциты;
• аутореактивные клетки.
Естественными факторами, инициирующими апоптоз, яляются глюкокортикоидные гормоны, Fas-лиганд, α-ФНО и другие иммуноцитокины, гранзимы и гранулизин. Апоптотическое уничтожение клеток-мишеней могут активировать Т-киллеры, ЕК с фенотипом CD16-CD56много и Т1-хелперы.
Помимо апоптоза возможен антителозависимый лимфоцитолиз. Например, с медицинской целью применяют антилимфоцитарную
сыворотку, которая в присутствии комплемента вызывает лизис лимфоцитов. Устранить лимфоидную популяцию возможно также воздействием ионизирующего излучения или цитостатиков.
Функциональная активность иммунокомпетентных клеток может быть ингибирована растворимыми факторами их конкурентов или потомков. Ведущая роль принадлежит иммуноцитокинам с множественными эффектами. Известно, например, что Т2- хелперы, γδТ-лимфоциты и тучные клетки при помощи ИЛ-4, 13 препятствуют дифференцировке Т0-хелпера в Т1-клетку. Последний, в свою очередь, может блокировать образование Т2-хелпера, синтезируя γ-ИФН. Пролиферацию Т- и В-лимфоцитов ограничивает β-ТФР, который продуцируют терминально дифференцированные Т-хелперы. Уже упомянутые продукты Т2-хелпера (ИЛ-4, 13 и β-ТФР) подавляют биологическую активность макрофагов.
Супрессия гуморального звена иммунитета может быть вызвана иммуноглобулинами. Избыточные концентрации иммуноглобулина класса G, связываясь со специальными рецепторами на мембране В-лимфоцита, тормозят биологическую активность клетки и ее способность дифференцироваться в плазмоцит.
|
|
Устранение из организма антигена в природе наблюдается при полном освобождении организма от патогена при развитии стерильного иммунитета. В клинической практике эффект достигается очищением организма плазмоили лимфосорбцией, а также нейтрализацией антигена антителами, специфичными для высокоиммуногенных эпитопов.
10.2.2.4. Возрастные изменения иммунной системы
В развитии иммунной системы четко прослеживаются два этапа. Первый, антигеннезависимый, который начинается с эмбрионального периода развития и частично продолжается всю жизнь. В течение этого периода образуются стволовые клетки и разнообразные антигенспецифические клоны лимфоцитов. Предшественники γδT и В1-лимфоцитов мигрируют в покровные ткани и формируют автономные лимфоидные ростки.
Второй этап, антигензависимый, продолжается с момента рождения особи до ее гибели. В этот период идет «ознакомление» иммунной системы с многообразием окружающих нас антигенов. По мере накопления биологического опыта, т.е. количества и качества продуктивных контактов с антигенами, происходят селекция
и тиражирование отдельных клонов иммунокомпетентных клеток. Особенно интенсивная экспансия клонов характерна для детского возраста. В течение первых 5 лет жизни иммунной системе ребенка приходится усваивать примерно 90% биологической информации. Еще 9% воспринимается до наступления пубертата, на взрослое состояние остается лишь около 1%.
Иммунной системе ребенка приходится справляться с чудовищными нагрузками, которые в основном падают на гуморальное звено иммунитета. В местах с повышенной плотностью населения и частыми межиндивидуальными контактами (крупные города) создаются условия для длительной персистенции высокой концентрации разнообразных патогенов. Поэтому дети в мегаполисах часто болеют. Однако создается впечатление о тотальном иммунодефиците, порожденном крайним экологическим неблагополучием. Между тем эволюционно заложенные механизмы иммунной защиты позволяют организму ребенка успешно справиться с трудными естественными испытаниями на жизнеспособность и адекватно отреагировать на вакцинопрофилактику.
|
|
С возрастом иммунная система меняет свою структуру. В организме взрослого до 50% всего лимфоидного пула представлено клонами клеток, прошедших антигенную стимуляцию. Накопленный иммунной системой биологический опыт проявляется образованием узкой «библиотеки» жизненно важных (актуальных) клонов лимфоцитов, специфичных для основных патогенов. Благодаря долгоживучести клеток иммунологической памяти актуальные клоны со временем становятся самодостаточными. Они приобретают способность к самоподдержанию и независимость от центральных органов иммунной системы. Функциональная нагрузка на тимус снижается, что проявляется его возрастной инволюцией. Тем не менее в организме сохраняется широкий набор невостребованных «наивных» клеток. Они способны отреагировать на любую новую антигенную агрессию.
Структура популяции Т-лимфоцитов также претерпевает возрастные изменения. Установлено, что в организме новорожденных преобладают Т2-хелперы, необходимые для развития антительной защиты. Однако со временем перед организмом все острее встает проблема внутриклеточного паразитизма, различных инвазий, мутаций, что требует надежного и хорошо организованного иммунологического надзора за морфогенетическим постоянством кле-
точных элементов организма. Поэтому после рождения начинает усиленно развиваться система адаптивного клеточного иммунитета, а вместе с ним образование клонов Т1-хелперов и Т-киллеров. Отмечено, что нарушение постнатальной колонизации желудочнокишечного тракта нормальной флорой тормозит процесс адекватного формирования популяции Т1-хелперов в пользу Т2-клеток. Избыточная активность последних оборачивается аллергизацией детских организмов.
Продуктивный иммунный ответ после своего завершения (нейтрализации и элиминации антигена из организма) также сопровождается изменениями клональной структуры антигенореактивных лимфоцитов. При отсутствии активирующих стимулов клон инволюционирует. Невостребованные клетки со временем погибают от старости или индукции апоптоза, причем этот процесс начинается с более дифференцированных лимфоцитов-эффекторов. Численность клона постепенно снижается и проявляется постепенным угасанием иммунного ответа. Однако в организме длительно персистируют клетки иммунологической памяти.
|
|
Старческий период жизни характеризуется доминированием в иммунной системе актуальных клонов антигенспецифических лимфоцитов в сочетании с нарастающей иммунодепрессией и снижением общей реактивности. Инфекции, вызванные даже условно-патогенными микробами, зачастую принимают затяжной или угрожающий характер. Клеточный иммунитет также теряет эффективность, постепенно нарастает объем злокачественно трансформированных клеток. Поэтому у пожилых людей часто встречаются новообразования.