

BKh_otvety_na_ekzamen_proshlogo_goda
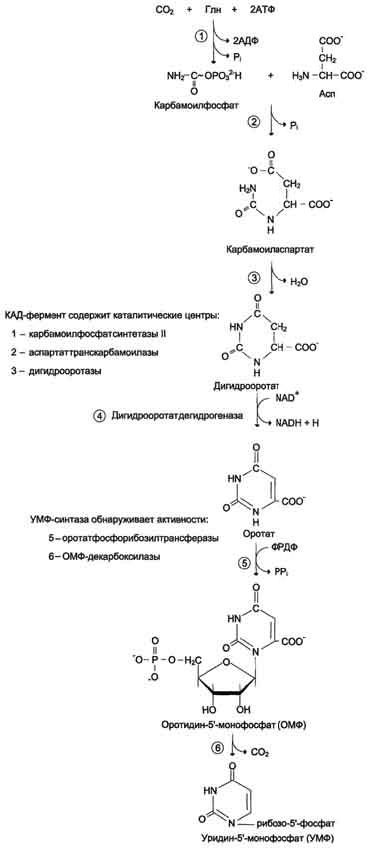
.pdfучастием КФС I. Карбамоилфосфат, использующийся на образование пирймидиновых нуклеотидов, является продуктом полифункционального фермента, который наряду с активностью КФС II содержит каталитические центры аспартаттранскарбамоилазы и дигидрооротазы. Этот фермент назвали "КАД-фермент" - по начальным буквам ферментативных активностей, которыми обладают отдельные каталитические домены этого белка. Объединение первых трёх ферментов метаболического пути в единый полифункциональный комплекс позволяет использовать почти весь синтезированный в первой реакции карбамоилфос-фат на взаимодействие с аспартатом и образование карбамоиласпартата, от которого отщепляется вода и образуется циклический продукт – дигидрооротат Отщепляясь от КАД-фермента, дигидрооротат подвергается дегидрированию NAD-зависимой дигидрооротатдегидрогеназой и превращается в свободное пиримидиновое основание - оротовую кислоту, или оротат.
 .
.
Образование УМФ. В цитозоле оротат становится субстратом бифункционального фермента - УМФ-синтазы, которая обнаруживает оротатфосфорибозилтранс-феразную и ОМФ-декарбоксилазную активности. Первоначально фосфорибозильный остаток от ФРДФ переносится на оротат и образуется нук-леотид -
оротидин-5'-монофосфат (ОМФ), декарбоксилирование которого даёт уридин-5-монофосфат (УМФ). Таким образом, шесть последовательных реакций синтеза пиримидиновых нуклеотидов осуществляются тремя ферментами, которые кодируются в геноме человека тремя различными структурными генами.
Биосинтез УДФ, УТФ и иитидиловых нуклеотидов УМФ под действием специфических нуклео-
зидмонофосфат (НМФ) и нуклеозиддифосфат (НДФ) киназ превращается в УДФ и УТФ в результате переноса γ-фосфатного остатка АТФ на соответствующий субстрат. НМФ-киназа катализирует следующую реакцию:
УМФ + АТФ |
УДФ + АДФ, |
а НДФ-киназа: |
|
УДФ + АТФ |
УТФ + АДФ. |
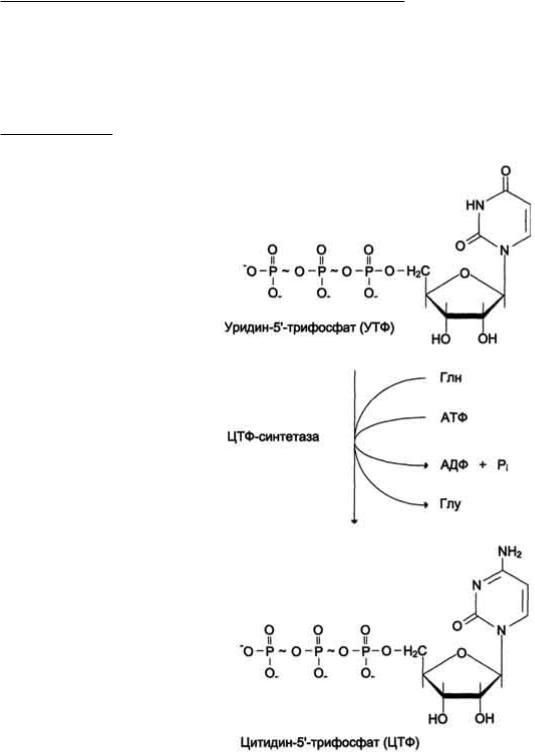
ЦТФ синтетаза катализирует амидирование УТФ, осуществляя АТФ-зависимое замещение кетогругшы урацила на амидную группу глутамина с образованием цитидин-5'-трифосфата (ЦТФ).
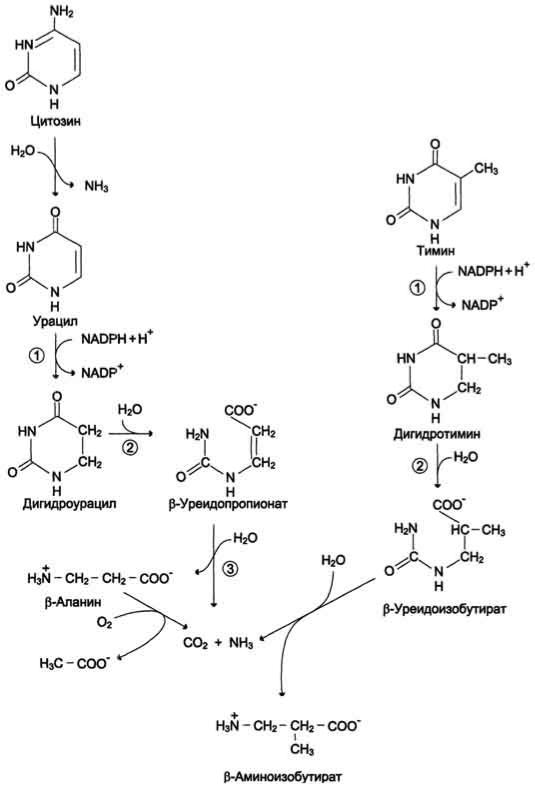
Пиримидиновые основания при участии дигидропиримидиндегидрогеназы присоединяют 2 атома водорода по двойной связи кольца с образованием дигидроурацила или дигидротимина. Оба гетероцикла могут взаимодействовать с водой в реакции, катализируемой дигидропиримидинциклогидролазой, и дигидроурацил превращается в β-уреидопропионовую кислоту, а дигидротимин - в β-уреидоизомасляную кислоту. Оба β-уреидопроизводных под действием общего для них фермента уреидопропионазы расщепляются с образованием СО2, NH4+ и β-аланина или β-аминоизомасляной кислоты соответственно. β- Аланин обнаруживают в плазме крови и многих тканях. Он используется в мышцах на образование дипептидов: карнозина и анзерина. Под действием бактериальной микрофлоры кишечника β-аланин включается в пантотеновую кислоту, которая всасывается и используется на образование КоА. Часть β- аланина и β-аминбутирата трансами-нируется с α-кетоглутаратом и даёт малонил полуальдегид или метилмалонил полуальдегид соответственно, которые превращаются в малонил-КоА и сукцинил-КоА и используются в соответствующих метаболических путях, либо окисляются до СО2 и Н2О. Частично β- аминобутират экскретируется с мочой.
101. Нарушения обмена нуклеотидов. Подагра; применение аллопуринола для лечения подагры. Ксантинурия. Оротацидурия.
Гиперурикемия подагры. Когда в плазме крови концентрация мочевой кислоты превышает норму, то возникает гиперурикемия. Вследствие гиперурикемии может развиться подагра - заболевание, при котором кристаллы мочевой кислоты и уратов откладываются в суставных хрящах, синовиальной оболочке, подкожной клетчатке с образованием подагрических узлов, или тофусов. К характерным признакам подагры относят повторяющиеся приступы острого воспаления суставов (чаще всего мелких) - так называемого острого подагрического артрита. Заболевание может прогрессировать в хронический подагрический артрит. Поскольку лейкоциты фагоцитируют кристаллы уратов, то причиной воспаления является разрушение лизосомальных мембран лейкоцитов кристаллами мочевой кислоты. Освободившиеся лизосомальные ферменты выходят в иитозоль и разрушают клетки, а продукты клеточного катаболизма вызывают воспаление.
Общий фонд сывороточных уратов в норме составляет ~ 1,2 г у мужчин и 0,6 г у женщин. При подагре без образования тофусов (т.е. подагрических узлов, в которых накапливаются ураты натрия и мочевая кислота) количество уратов возрастает до 2-4 г, а у пациентов с тяжёлой формой болезни, сопровождающейся ростом тофусов, может достигать 30 г. Подагра - распространённое заболевание, в разных странах ею страдают от 0,3 до 1,7% населения. А поскольку сывороточный фонд уратов у мужчин в 2 раза больше, чем у женщин, то они и болеют в 20 раз чаще, чем женщины. Как правило, подагра генетически детерминирована и носит семейный характер. Она вызвана нарушениями в работе ФРДФ синтетазы или ферментов "запасного" пути: гипоксантин-гуанин- или аденинфосфорибозилтрансфераз. К другим характерным проявлениям подагры относят нефропатию, при которой наблюдают образование уратных камней в мочевыводящих путях.
Полиморфные варианты ФРДФ синтетазы. Активность ФРДФ синтетазы, катализирующей образование ФРДФ, строго контролируется пуриновыми нуклеотидами. Мутации в гене ФРДФ синтетазы привели к появлению полиморфных вариантов фермента, которые характеризуются аномальным ответом на обычные регуляторные факторы: концентрацию рибозо-5-фосфата и пуриннуклеотидов. Как правило, наблюдаетсясуперактивация фермента. Пуриновые нуклеотиды синтезируются со скоростью, почти независимой от нужд клетки. Это вызывает ингибирование запасных "путей спасения", усиление катаболизма избыточного количества нуклеотидов, повышение продукции мочевой кислоты, гиперурикемию и подагру. Примерно у 40% больных одной из форм гликогеноза - болезнью Гирке (недостаточностью глюкозо-6-фосфатазы) сопутствующей патологией является подагра. Снижение способности печени секретировать глюкозу в кровь увеличивает использование глюкозо-6-фосфата в пентозофосфатном пути. Образуются большие количества рибозо-5-фосфата, которые могут стимулировать избыточный синтез, а следовательно, и катаболизм пуриновых нуклеотидов.
Дефектный |
Характер дефекта |
Клинические проявления |
Заболевание |
фермент |
|
|
|
|
|
|
|
ФРДФ синтетаза |
Суперактивация и Vmax |
Гиперурикемия, повышенная экскреция |
Подагра |
|
Устойчивость к |
уратов с мочой, подагрический артрит |
|
|
ретроингибированию |
|
|
|
Снижение Кm для |
|
|
|
рибозо-5-фосфата |
|
|
|
|
|
|
Гипоксантин- |
Частичная потеря |
Те же |
Подагра |
гуанинфосфори- |
активности |
|
|
бозилтрансфераз |
|
|
|
а |
|
|
|
|
Полная потеря |
Гиперурикемия, нефропатия, артрит, |
Синдром Лёша- |
|
активности |
неврологические и психические |
Нихена |
|
|
отклонения |
|
|
|
|
|
Аденинфосфориб |
Полная потеря |
Образование камней 2,8- |
Почечнокаменная |
о- |
активности |
дигидроксиаденина |
болезнь |
зилтрансфераза |
|
|
|
|
|
|
|
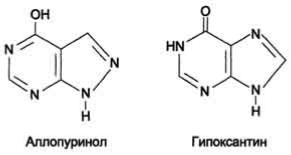
Лечение гиперурикемииОсновным препаратом, используемым для лечения гиперурикемии, является аллопуринол - структурный аналог гипоксантина.
Аллопуринол оказывает двоякое действие на обмен пуриновых нуклеотидов:
•ингибирует ксантиноксидазу и останавливает катаболизм пуринов на стадии образования гипоксантина, растворимость которого почти в 10 раз выше, чем мочевой кислоты. Действие препарата на фермент объясняется тем, что сначала он, подобно гипоксанти-ну, окисляется в гидроксипуринол, но при этом остаётся прочно связанным с активным центром фермента, вызывая его инактивацию;
•с другой стороны, будучи псевдосубстратом, аллопуринол может превращаться в нуклеотид по "запасному" пути и ингибировать ФРДФ синтетазу и амидофосфорибозилтрансферазу, вызывая торможение синтеза пуринов de novo.
При лечении аллопуринолом детей с синдромом Лёша-Нихена удаётся предотвратить развитие патологических изменений в суставах и почках, вызванных гиперпродукцией мочевой кислоты, но препарат не излечивает аномалии в поведении, неврологические и психические расстройства.
Оротацидурия. Это единственное нарушение синтеза пиримидинов de novo. Оно вызвано снижением активности УМФ-синтазы, которая катализирует образование и декарбоксилирование ОМФ. Поскольку в эмбриогенезе от образования пиримидинов de novo зависит обеспечение синтеза ДНК субстратами, то жизнь плода невозможна при полном отсутствии активности этого фермента. Действительно, у всех пациентов с оротацидурией отмечают заметную, хотя и очень низкую активность УМФ-синтазы. Установлено, что содержание оротовои кислоты в моче пациентов (1 г/сут и более) значительно превосходит количество оротата, которое ежедневно синтезируется в норме (около 600 мг/сут). Снижение синтеза пиримидиновых нуклеотидов, наблюдающееся при этой патологии, нарушает регуляцию КАД-фермента по механизму ретроингибирования, из-за чего возникает гиперпродукция оротата. Клинически наиболее характерное следствие оротацидурии - мегалобластная анемия, вызванная неспособностью организма обеспечить нормальную скорость деления клеток эритроцитарного ряда. Её диагностируют у детей на том основании, что она не поддаётся лечению препаратами фолиевой кислоты. Недостаточность синтеза пиримидиновых нуклеотидов сказывается на интеллектуальном развитии, двигательной способности и сопровождается нарушениями работы сердца и ЖКТ. Нарушается формирование иммунной системы, и наблюдается повышенная чувствительность к различным инфекциям. Гиперэкскреция оротовои кислоты сопровождается нарушениями со стороны мочевыводя-щей системы и образованием камней. При отсутствии лечения больные обычно погибают в первые годы жизни. При этом оротовая кислота не оказывает токсического эффекта. Многочисленные нарушения в работе разных систем организма вызваны "пиримидиновым голодом". Для лечения этой болезни применяют уридин (от 0,5 до 1 г/сут), который по "запасному" пути превращается в УМФ.
Уридин + АТФ УМФ + АДФ.
Нагрузка уридином устраняет "пиримидиновый голод", а поскольку из УМФ могут синтезироваться все остальные нуклеотиды пиримидинового ряда, то снижается выделение оротовои кислоты из-за восстановления механизма ретроингибирования КАД-фермента. Для больных оротацидурией лечение уридином продолжается в течение всей жизни, и этот нуклеозид становится для них незаменимым пищевым фактором.
Кроме генетически обусловленных причин, оротацидурия может наблюдаться:
•при гипераммониемии, вызванной дефектом любого из ферментов орнитинового цикла,
•за исключением карбамоилфосфатсинтетазы I. В этом случае карбамоилфосфат, синтезированный в митохондриях, выходит в цитозоль клеток и начинает использоваться на образование пиримидиновых нуклеотидов. Концентрация всех метаболитов, в том числе и оротовой кислоты, повышается.
Наиболее значительная экскреция оротата отмечается при недостаточности орнитинкарбамоилтрансферазы (второго фермента орнитинового цикла);
•в процессе лечения подагры аллопуринолом, который превращается в оксипуринолмононуклеотид и становится сильным ингибитором УМФ-синтазы. Это приводит к накоплению оротовой кислоты в тканях и крови.
Нарушения катаболизма пиримидинов
Известны нарушения в работе 2 ферментов этого метаболического пути. При недостаточности пиримидин-5'- нуклеотидазы нарушаются отщепление неорганического фосфата от пиримидиновых мононуклеотидов и образование нуклеозидов. Неактивная изоформа пиримидин-5'-нуклеотидазы обнаружена в эритроцитах. В результате наблюдается накопление гиримидиновых НТФ, которые ингибируют пентозофосфатный путь п р е в р а щ е н и я г л ю к о з ы и т е м с а м ы м с о з д а ю т п р е д п о с ы л к и к г е м о л и з у э р и т р о ц и т о в . Дигидропиримидиндегидрогеназа - скорость-лимитирующий фермент катаболизма пиримидинов.
Нарушение работы этого фермента сопровождается отклонениями в функционировании нервной системы и диагностируется на основании повышения уровня свободных пиримидинов: урацила и тимина в плазме крови.
102. Биосинтез дезоксирибонуклеотидов. Применение ингибиторов синтеза дезоксирибонуклеотидов для лечения злокачественных опухолей.
Синтез дезоксирибонуклеотидов идёт с заметной скоростью только в тех клетках, которые вступают в S- фазу клеточного цикла и готовятся к синтезу ДНК и делению. В покоящихся клетках дезоксинуклеотиды практически отсутствуют. Все дезоксинуклеотиды, кроме тимидиловых, образуются из рибонуклеотидов путём прямого восстановления ОН-группы у второго углеродного атома рибозы в составе рибонуклеозиддифосфатов до дезоксирибозы. Тимидиловые нук-леотиды синтезируются из dУМФ особым путём с участием N5,N10-метилен-Н4-фолата.
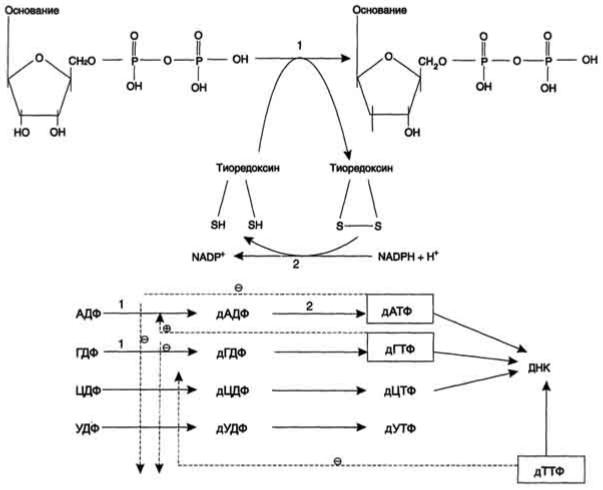
Рибонуклеотидредуктазный комплекс. Реакцию восстановления НДФ в дезоксипроизводные катализирует рибонуклеотидредук-тазный комплекс, в состав которого входят: собственно рибонуклеотидредуктаза (РНР), белок тиоредоксин и фермент тиоредоксинредуктаза, обеспечивающий регенерацию восстановленной формы тиоредоксина.
Рибонуклеотидредуктаза - олигомерный белок, состоящий из двух В1- и двух В2-субъединиц, и содержит негеминовое железо в качестве кофактора. Непосредственным донором водорода в реакции восстановления рибозы служит низкомолекулярный белок тиоредоксин. В рабочую часть этого белка входят 2 SH-группы, которые, отдавая водород, окисляются с образованием дисульфидного мостика. Второй фермент комплекса - тиоредоксинредуктаза - катализирует гидрирование окисленного тиоредоксина с использованием NADPH. При участии комплекса РНР образуются: dАДФ, dГДФ, dУДФ и dЦДФ, которые с помощью НДФ-киназ превращаются в дНТФ, 3 из которых (кроме дУДФ) непосредственно используются в синтезе ДНК.
дНДФ + АТФ дНТФ + АДФ.
Биосинтез тимидиловых нуклеотидов Тимидин-5'-монофосфат (дТМФ) образуется из дУМФ в реакции, катализируемой тимиди-латсинтазой (рис. 10-18). Донором метильной группы, появляющейся в 5-положении пиримидинового кольца в молекуле дТМФ, служит кофермент тимидилатсинтазы - N5,N10-метилен-Н4-фолат. С помощью этого кофермента в молекулу дУМФ включается метиленовая группа и восстанавливается в метальную, используя 2 атома водорода от Н4-фолата. Образование субстрата тимидилатсинтазной реакции - дУМФ осуществляется двумя путями:
•дефосфорилированием дУДФ;
•гидролитическим дезаминированием дЦМФ с помощью дЦМФ дезаминазы. дЦМФ получается при дефосфорилировании дЦДФ - одного из продуктов рибонуклеотидредуктаз-ной реакции. В организме человека это основной путь образования дУМФ.
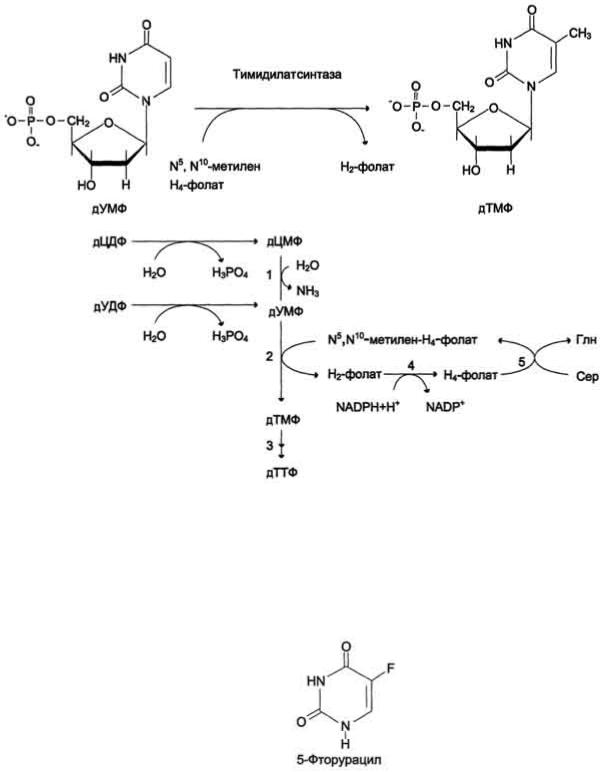
Скорость синтеза дТМФ зависит также от количества второго субстрата тимидилатсинтазной реакции - N5,N10-метилен-Н4-фолата, пополнение запасов которого осуществляется при участии 2 ферментов: дигидрофолатредуктазы, которая с участием NADPH восстанавливает Н 2-фолат в Н 4-фолат, и серии гидроксиметилтрансферазы, осуществляющей перенос β-гидроксиметиленовой группы серина на Н 4-фолат. У человека дТМФ образуется, главным образом, из дЦДФ.
Противоопухолевые препараты. Синтезировано очень много аналогов дНТФ, которые включаются ДНК полимеразами в ДНК и ингибируют репликацию. К числу мощных противоопухолевых препаратов принадлежит 5-фторурацил (5-FU) - аналог урацила.
В организме по "запасным" путям 5-FU превращается в 5-Р-УМФ либо в реакции, катализируемой оротатфосфорибозилтрансферазой, либо через промежуточное образование нуклеозида и последующее фосфорилирование. Превращаясь в нуклеозиддифосфат, 5-FU может участвовать в реакции, катализируемой РНР, и восстанавливаться в соответствующее дезоксипроизводное. Под действием фосфатазы 5-Р-дУДФ снова теряет фосфат, и образующийся 5-Р-дУМФ связывается с тимидилатсинтазой и N5, N10-метилен-Н4-фолатом, образуя комплекс, напоминающий промежуточное соединение в реакции превращения дУМФ в дТМФ. Тимидилатсинтаза оказывается полностью блокированной, и синтез дТМФ прекращается.
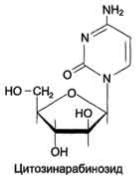
Цитозинарабинозид (или цитарабин) представляет собой соединение, в котором остаток ри-бозы замещён на стериоизомер - арабинозу. Оно используется в химиотерапии рака, в частности, при острой миелоцитарной лейкемии. В организме препарат может превращаться в дНТФ, ингибировать ДНК полимеразы и снижать скорость репликации.
103. Биосинтез ДНК, субстраты, источники энергии, матрица, ферменты. Понятие о репликативном комплексе. Этапы репликации.
Этапы биосинтеза ДНК. Предложен ряд моделей механизма биосинтеза ДНК с участием указанных ранее ферментов и белковых факторов, однако детали некоторых этапов этого синтеза еще не выяснены. Основываясь главным образом на данных, полученных в опытах in vitro, предполагают, что условно
механизм синтеза ДНК у Е. coli может быть подразделен на три этапа; инициацию, т.е. начало, элонгацию, т.е. продолжение, и терми-нацию, т.е. завершение (прекращение) синтеза. Каждый из этих этапов требует участия специфических ферментов и белковых факторов.
Этап I – инициация биосинтеза ДНК – в определённом сайте (точка начала репликации) происходит локальная денатурация ДНК, цепи расходятся и образуются две репликативные вилки, движущиеся в противоположных направлениях. В образовании репликативной вилки принимает участие ряд белков и ферментов. Так, семейство ДНК-топоизомераз (I, II и III), обладая нуклеазной активностью, участвует в регуляции суперспирализации ДНК. Например, ДНК-топоизомераза I разрывает фосфоэфирную связь в одной из цепей двойной спирали и ковалентно присоединяется к 5'-концу в точке разрыва . По окончании формирования репликативной вилки фермент ликвидирует разрыв в цепи и отделяется от ДНК. Разрыв водородных связей в двухцепочечной молекуле ДНК осуществляет ДНК-хеликаза.Фермент ДНК-хеликаза использует энергию АТФ для расплетения двойной спирали ДНК. В результате происходит раскручивание участка суперспирализованной молекулы ДНК. В поддержании этого участка ДНК в раскрученном состоянии участвуют SSB-белки (от англ, single strand binding proteins, т.е. белки, связывающиеся с одноцепочечными нитями ДНК). SSB-белки, не закрывая азотистых оснований, связываются с одноцепочечной ДНК по всей длине разделившихся цепей и таким образом предотвращают их комплементарное скручивание и образование "шпилек". Они обладают большим сродством к одноцепочечным участкам ДНК, независимо от первичной структуры цепей.
Этап II – элонгация синтеза ДНК – репликация ДНК осуществляется ДНК-зависимыми ДНК-полимеразами (рис. 4-16). Субстратами и источниками энергии для синтеза продукта служат 4 макроэргических соединения - дезоксирибонуклеозидтрифосфаты дАТФ, дГТФ, дЦТФ и дТТФ, для активации которых необходимы ионы магния. Нейтрализуя отрицательный заряд нуклеотидов, они повышают их реакционную способность. Ферменты проявляют каталитическую активность только в присутствии предварительно
раскрученной матричной двухцепочечной ДНК. Синтез цепей ДНК происходит в направлении 5' 3' растущей
цепи, т.е. очередной нуклеотид присоединяется к свободному 3'-ОН-концу предшествующего нуклеотид-ного остатка. Синтезируемая цепь всегда антипараллельна матричной цепи. В ходе репликации образуются 2 дочерние цепи, представляющие собой копии матричных цепей. В синтезе эукариотических ДНК принимают участие 5 ДНК-полимераз (α, β, γ, δ, ε). ДНК-полимеразы различают по числу субъединиц, молекулярной массе, ассоциации с разными вспомогательными белками, ускоряющими процесс биосинтеза ДНК, и функциональному назначению. ДНК-полимеразы α (альфа), β (бета), δ (дельта), ε (эпсилон) участвуют в синтезе ДНК в ядре клеток, ДНК-полимераза γ (гамма) - в репликации митохондриальной ДНК. ДНКполимеразы β, δ, ε не могут инициировать образование дочерних цепей, так как не имеют сродства к одиночной нити ДНК. Инициирует репликацию ДНК-полимераза α, которая комплементарна определённому сайту одноцепо-чечной ДНК. Присоединяясь к нему, ДНК-полимераза а синтезирует небольшой фрагмент РНК - праймер, состоящий из 8-10 рибонуклеотидов. ДНК-полимераза а состоит из четырёх субъединиц. Каждая из субъединиц фермента выполняет определённую функцию: "узнавание" сайта репликации, синтез праймера (8-10 рибонуклеотидов), синтез фрагмента цепи ДНК, около 50 дезоксирибонуклеотидов. Таким образом, ДНК-полимераза α синтезирует олигонуклеотид, содержащий примерно 60 нуклеотидных остатков; первые 8-10 представлены рибонуклеотидами (праймер), а остальные - дезоксирибонуклеотидами.
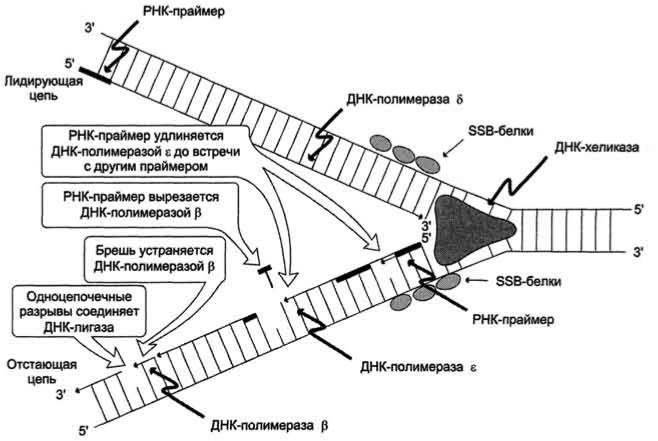
ДНК-полимераза δ
Олигонуклеотид, синтезированный ДНК-полимеразой α и образующий небольшой двухцепочечный фрагмент с матрицей, позволяет присоединиться ДНК-полимеразе δ и продолжить синтез новой цепи в направлении от 5'- к 3'-концу по ходу раскручивания репликативной вилки. ДНК-полимераза δ последовательно наращивает цепь, шаг за шагом присоединяя к ней соответствующие дезоксинуклеотиды. Выбор ДНКполимеразой δ очередного нуклеотида определяется матрицей. Включение дезоксирибонуклеозидмонофосфатов в растущую цепь ДНК сопровождается гидролизом макроэргических связей соответствующих нуклеозидтрифосфатов и отщеплением пирофосфата (Н4Р2О7). Энергия макроэргических связей расходуется на образование 3',5'-фосфодиэфирной связи между последним нуклеотидом растущей цепи ДНК и присоединяемым нуклеотидом. Включение нуклеотида в синтезируемую цепь ДНК невозможно без предварительного связывания азотистого основания водородными связями с комплементарным нуклеотидом матричной цепи. ДНК-полимеразы (α, β, γ, δ, ε) могут синтезировать
нуклеотидную цепь только в направлении 5' 3', матричная цепь всегда считывается в направлении 3' 5'. В
каждой репликативной вилке идёт одновременно синтез двух новых (дочерних) цепей. Направление синтеза цепи ДНК совпадает с направлением движения репликативной вилки лишь для одной из вновь синтезируемых цепей (лидирующая цепь). На второй матричной цепи синтез дочерней ДНК осуществляется
двумя ферментами: ДНК-полимеразой α и ДНК-полимеразой ε в направлении 5' 3', но против движения
репликативной вилки. Поэтому вторая цепь синтезируется прерывисто, короткими фрагментами, которые называют "фрагменты Оказаки" (по имени открывшего их исследователя). Дочерняя цепь ДНК, синтез которой происходит фрагментами, называют отстающей цепью. Каждый фрагмент Оказаки, примерно 100 нуклеотидных остатков, содержит праймер. Праймеры удаляет ДНК-полимераза β, постепенно отщепляя с 3'-конца фрагмента по одному ри-бонуклеотиду. К ОН-группе на 3'-конце предыдущего фрагмента ДНКполимераза β присоединяет дезоксирибонуклеотиды в количестве, равном вырезанному праймеру и таким образом заполняет брешь, возникающую при удалении рибонуклеотидов. Фермент ДНК-лигаза катализирует образование фосфодиэфирной связи между 3'-ОН-группой дезоксирибозы одного фрагмента цепи ДНК и 5'- фосфатом следующего фрагмента. Реакция протекает с затратой энергии АТФ. Таким образом, из множества фрагментов Оказаки образуется непрерывная цепь ДНК.
Этап III – терминация синтеза ДНК – наступает, скорее всего, когда исчерпана ДНК-матрица и трансферазные реакции прекращаются. Точность репликации ДНК чрезвычайно высока, возможна одна ошибка на 1010 трансферазных реакций, однако подобная ошибка обычно легко исправляется за счет процессов репарации
104. Синтез ДНК и фазы клеточного деления. Роль циклинов и циклинзависимых протеиназ в продвижении клетки по клеточному циклу.
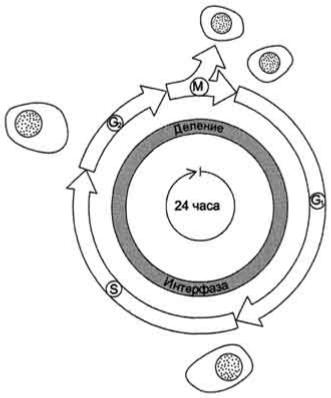
Процессы роста и деления клеток лежат в основе жизни любого организма. Но прежде чем совершить деление, клетка должна с высокой точностью копировать свой геном, синтезировать множество высоко- и низкомолекулярных соединений. Совокупность событий, обеспечивающих деление эукариотических клеток, называют "клеточный цикл" . Продолжительность клеточного цикла зависит от типа делящихся клеток, у взрослого человека она может варьировать примерно от 8 ч и более, а для некоторых типов клеток до года и больше.
Все фазы клеточного цикла G 1, S, G2, M могут различаться по длительности, но в особенности это касается фазы G 1, длительность которой может быть равна практически нулю или быть столь продолжительной, что может казаться, будто клетки вообще прекратили деление. В этом случае говорят, что клетки находятся в состоянии покоя (фаза G 0). Так, нейроны взрослого человека не делятся вообще. Клетки эпителия кишечника делятся на протяжении всей жизни человека, но даже у этих быстропролиферирующих клеток подготовка к делению занимает 24 ч. Клетки лёгких, почек, печени во взрослом организме начинают делиться только лишь в ответ на повреждение органов. Внешние сигналы могут стимулировать или ингибировать прохождение клетки через цикл. Пролиферативные сигналы очень разнообразны, они зависят от типа клетки, стадии развития и других факторов. Такими сигналами могут быть факторы роста, интерлейкины, гормоны, способные поддерживать или индуцировать пролиферацию определённых типов клеток. Сигнальные молекулы связываются специфическими мембранными рецепторами, активируют внутриклеточные пути передачи сигналов от рецептора к ядру и таким образом индуцируют транскрипцию определённых генов. Одними из первых активируются гены, кодирующие белки циклины.
Циклин |
Киназа |
Функция |
|
|
|
D,E |
CDK4, CDK6 |
Регулирует переход клетки из G1-фазы в S-фазу |
|
|
|
А |
CDK2 |
Активирует синтез ДНК на начальной стадии S-фазы |
|
|
|
В |
CDK1 |
Регулирует переход клетки из G2-фазы в М-фазу |
|
|
|
Белки были названы циклинами, потому что их концентрация в клетке периодически меняется по мере прохождения клеткой разных фаз клеточного цикла. Все циклины делят на 2 подсемейства: G1-циклины (D, Е) и митотические циклины (А и В). Любой из циклинов представлен группой полиморфных белков, например циклин D представлен формами Dl, D2, D3. У каждого типа циклинов есть гомологичный участок из 100 аминокислотных остатков - "циклиновый бокс", отвечающий за связывание с циклинзависимой киназой (от англ. CDK - cyclin-dependent kinases). В клетках эукариотов существует примерно восемь различных CD К (CDK1-8), активирующихся различными циклинами. Циклинзависимые киназы, связывая циклин, переходят в активную форму и могут фосфорилировать специфические белки, например факторы транскрипции, белки-ингибиторы факторов транскрипции, которые регулируют синтез ферментов, обеспечивающих репликацию. Синтез каждого циклина начинается при подготовке к соответствующей фазе клеточного цикла, его концентрация в клетке повышается, а после окончания фазы резко падает до нуля. Завершившие свою работу комплексы циклинов и CDK связываются специфическими белками, ингибирующими их активность, и затем подвергаются разрушению.