

Классификация моносахаридов
Не все следующие монозы найдены в природе, некоторые были синтезированы:
|
Альдоза | ||||
|
Альдотриоза |
| |||
|
Альдотетрозы |
|
| ||
|
Альдопентозы |
|
|
|
|
 глицеральдегид
(глицераль)
глицеральдегид
(глицераль) эритроза
эритроза треоза
треоза рибоза
рибоза арабиноза
арабиноза ксилоза
ксилоза ликсоза
ликсоза
|
Альдогексозы |
аллоза |
альтроза |
глюкоза |
манноза | ||
|
гулоза |
идоза
|
галактоза |
талоза | |||
|
Кетозы |
| |||||
|
кетотриоза |
|
| ||||
|
кетотетрозы |
|
| ||||
|
кетопентозы |
|
|
| |||
|
кетогексозы |
|
|
|
|
| |








 эритрулоза
эритрулоза рибулоза
рибулоза ксилулоза
ксилулоза псикоза
псикоза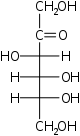 фруктоза
фруктоза сорбоза
сорбоза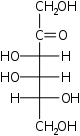 тагатоза
тагатоза


В природе широко распространены гексозы (D-глюкоза, D-галактоза, D-манноза, D-фруктоза) и пентозы (D-рибоза, D-ксилоза, D-арабиноза). Среди производных моносахаридов наиболее распространенными являются аминосахара D-глюкозамин и D-галактозамин и дезоксисахар 2-дезокси-D-рибоза.

Моносахариды с более длинными цепями известны, например кетогепсозы,маногептулозаиседогептулоза.
Химические свойства
Моносахариды вступают в химические реакции, свойственные карбонильнойигидроксильнойгруппам. Характерная особенность моносахаридов — способность существовать в открытой (ациклической) и циклической формах и давать производные каждой из форм. Большинство моноз циклизуются в водном растворе с образованиемгемиацеталейилигемикеталей(в зависимости от того, являются ли они альдозами или кетозами) между спиртом и карбонильной группой того же самого сахара. Глюкоза, например, легко образуетполуацетали, при взаимодействии гидроксильной группы возле пятого атома углерода с альдегидной группой (С1) образуется 6-членное кольцо, названноепиранозид. Та же самая реакция может иметь место между С1(альдегидной группой) и О4 (гидроксильной группы), чтобы сформировать 5-членноефуранозид.
Полезно подробно познакомиться с глюкозой – наиболее распространенным в природе моносахаридом. Глюкоза относится к гексозам; ее формула С6Н12O6. Моносахариды. Альдозы. Кетозы. Открытая цепь и циклические формы углеводов. Открытая цепь и циклические формы углеводов на рисунке молекула глюкозы представлена в виде «открытой цепи» и в виде циклической структуры. Открытая цепь может быть прямолинейной цепочкой, но углы связей между атомами углерода допускают у пентоз и гексоз образование стабильных циклических структур. У гексоз типа глюкозы первый атом углерода соединяется с кислородом при пятом углеродном атоме, что приводит к образованию шестичленного кольца. Обратите внимание на то, что атом кислорода включается в кольцо, а шестой углеродный атом оказывается вне кольца. У пентоз первый атом углерода соединяется с кислородом при четвертом углеродном атоме, в результате чего образуется пятичленное кольцо, как показано на рисунке. Циклические структуры пентоз и гексоз – обычные их формы; в каждый данный момент лишь небольшая часть молекул существует в виде открытой цепи. В состав дисахаридов и полисахаридов также входят циклические формы моносахаридов.
Альфа- и бета- изомеры углеводов. Циклические структуры могут существовать в двух формах, известных как альфа(α)- и бета(β) -формы. У α-формы гидроксильная группа при первом углеродном атоме расположена под плоскостью цикла, а у β-формы — над ней. Такие молекулы, с одинаковой химической формулой, но с разной структурой, называются изомерами. На рисунке представлены пространственные модели α - и β -изомеров глюкозы. В любой данный момент в растворе глюкозы часть молекул существует в виде открытой цепи и часть — в циклической форме. Последняя более стабильна и потому преобладает. Возможны спонтанные переходы от открытой цепи к одной из двух циклических форм, и наоборот. При этом в конечном счете неизменно достигается равновесие, при котором соотношение различных форм остается постоянным.

Как указано выше, только циклические структуры глюкозы образуют дисахариды и полисахариды. Хотя структурные различия между α - и бета-глюкозой невелики, но сказываются на свойствах образуемых ими молекул они весьма существенно. Ниже мы увидим, что α -глюкоза входит в состав крахмала, а бета-глюкоза – в состав целлюлозы, двух полисахаридов с весьма различными свойствами.
Стереоизомеры моносахаридов обладают оптической активностью: при прохождении через их растворы луча поляризованного света они отклоняют плоскость поляризации на некоторый характерный для каждого вещества угол вправо или влево. Этим свойством оптических стереоизомеров пользуются для количественного определения их содержания в биологических жидкостях. Наиболее просто устроенным моносахаридом. имеющим оптически активные стереоизомеры, является глицериновый альдегид. Он существует в двух формах: D (правовращающей) и L (левовращающей),
н
I
С=О
I
Н-С*-ОН
I
н-с-он
I
|
Н I С=О I Н-С*-ОН I Н-С-ОН I Н D-глицериновый альдегид |
Н I С=О I НО-С*-Н I Н-С-ОН I Н L-глицериновый альдегид |
Принадлежностъ моносахарида к D- или L-ряду определяется положением водорода и гидроксила у наиболее удаленного от альдегидной или кетонной группы асимметричного углеродного атома (он обозначен звездочкой).
Кроме того, принадлежностъ моносахарида к D- или L-ряду не определяет направление вращения плоскости поляризации.
Это направление обозначается дополнительно знаком (+) при правом вращении и знаком (‒) при левом вращении.
Олигосахариды(отгреч.ὀλίγος— немногий). состоят из нескольких моносахаридов, соединенных ковалентными связями. При гидролизе они распадаются на входящие в них моносахариды. Олигосахариды, состоящие из одинаковых моносахаридных остатков, называют гомополисахаридами, а из разных–гетерополисахаридами.
Примером наиболее распространённых олигосахаридов могут служить дисахариды и трисахариды. Дисахариды состоят из двух молекул моносахаридов. Наиболее распространенные дисахариды сахароза (пищевой или тростниковый сахар), состоящий из остатков глюкозы и фруктозы, лактоза(молочный сахар), состоящий из остатков глюкозы и галактозы.
Трисахариды отличаются одним от другого структурой входящих в них моносахаридов и их последовательностью в цепи. Так, например:
Рафиноза – состоит из остатков D-галактозы, D-глюкозы и D-фруктозы.
Мелицитоза – состоит из двух остатков D-глюкозы и одного остатка D-фруктозы.
Мальтотриоза – состоит из трех остатков D-мальтозы.
В зависимости от строения дисахариды делятся на две группы: восстанавливающие и невосстанавливающие.
Олигосахариды состоят из нескольких моносахаридов, соединенных ковалентными связями. При гидролизе они распадаются на входящие в них моносахариды. Примером олигосахаридов могут служить дисахариды, состоящие из двух молекул моносахаридов. Наиболее распространенные дисахариды сахароза (пищевой или тростниковый сахар), состоящий из остатков глюкозы и фруктозы, лактоза(молочный сахар), состоящий из остатков глюкозы и галактозы.
Полисахариды представляют собой длинные неразветвленные цепи. Включающие сотни и тысячи моносахаридных остатков. Наиболее известные из них – крахмал, целлюлоза, гликоген - состоят из остатков глюкозы.
Функции углеводов в организме весьма разнообразны.
1. Энергетическая.
2. Структурная функция (входят в состав клеточных структур).
3. Защитная (синтез иммунных тел в ответ на антигены).
4.Антисвертывающая (гепарин).
5. Гомеостатическая (поддержание водно-солевого обмена)
6. Механическая ( входят в состав соединительных и опорных тканей).
Превращения углеводов в пищеварительной системе
Гидролитическое расщепление углеводов в процессе пищеварения происходит под действием ферментов гликозидаз, расщепляющих 1-4 и 1-6-гликозидазные связи в молекулах сложных углеводов. Простые углеводы пищеварению не подвергаются, может только происходить брожение некоторой части их в толстом кишечнике под действием ферментов микроорганизмов.
К гликозидазамотносятсяамилазы слюны, поджелудочного и кишечного соков, мальтазы слюны и кишечного сока, конечная декстриназа, сахараза и лактаза кишечного сока. Гликозидазы активны в слабощелочной и угнетаются в кислой среде, за исключением амилазы слюны, которая катализирует гидролиз полисахаридов в слабокислой среде и теряет активность при увеличении кислотности.
В ротовой полости начинается пищеварение крахмала, на который действует амилаза слюны, расщепляющая 1-4-связи между глюкозными остатками внутри молекул амилозы и амилопектина и приводящая к образованию из них декстринов и мальтозы. В слюне содержится в небольших количествах имальтаза,гидролизующая мальтозу до глюкозы. Другие дисахариды пищи во рту не расщепляются.
Дальнейший распад декстринов, а также нерасщепленного крахмала и гликогена протекает в тонкой кишке с участием амилазы поджелудочного сока. В результате образуется дисахарид мальтоза, состоящая из двух остатков глюкозы. Завершается переваривание углеводов превращением образовавшейся мальтозы и других пищевых дисахаридов (сахароза, лактоза) в моносахариды (глюкоза, фруктоза, галактоза), главным из которых является глюкоза.
Клетчатка (целлюлоза) в молекуле которой остатки глюкозы соединены прочными связями, в ходе пищеварения не расщепляется и, пройдя через весь кишечник, выделяется из организма.
Образующаяся при пищеварении смесь моносахаридов, на 6О% состоящая из глюкозы, всасывается стенками тонкого кишечника, поступает в кровь и разносится к клеткам тела, в кишечной стенке фруктоза и галактоза частично превращаются в глюкозу.
Всасывание моносахаридов – активный физиологический процесс, протекающий с затратой энергии. Ее обеспечивают окислительные процессы, происходящие в клетках кишечной стенки. Мноосахариды получают энергию, взаимодействуя с АТФ в реакциях, продуктами которых являются фосфорные эфиры моносахаридов. При переходе из кишечной стенки в кровь фосфорные эфиры расщепляются фосфатазами, и в кровоток поступают свободные моносахариды. Поступление их из крови в клетки различных органов также сопровождается их фосфорилированием.
Образовавшиеся моносахариды всасываются по системе воротной вены и поступают вначале в печень. При этом в печень поступает практически только глюкоза, так как в ходе всасывания в клетках тонкой кишки в нее могут превращаться другие моносахариды (фруктоза, галактоза и др.).
В печени значительная часть глюкозы превращается в гликоген, который представляет собою запасную, резервную форму глюкозы или депо глюкозы (свободная глюкоза накапливаться в клетках не может, так как ее молекулы имеют малый размер и легко проходят через клеточные мембраны). Между приемами пищи в печени протекает противоположный процесс – гликоген распадается на глюкозу, которая из печени выходит в кровь.
На поверхности всех клеток имеются углеводы. Это полисахаридные и олигосахаридные цепи, ковалентно присоединенные к мембранным белкам и липидам. Углеводы всегда располагаются на той стороне мембраны, которая не контактирует с цитозолем. То есть, на внешних (плазматических) мембранах они присоединяются снаружи клетки. Функция углеводов клеточной поверхности пока неизвестна, но представляется вероятным, что некоторые из них принимают участие в процессах межклеточного узнавания.
Механизмы транспорта веществ через плазмолемму.
Плазматическая мембрана, или плазмалемма,представляет собой поверхностную периферическую структуру, ограничивающую клетку снаружи и обеспечивающую ее связь с другими клетками и внеклеточной средой. Она имеет толщину около 10 нм. Среди других клеточных мембран плазмалемма является самой толстой. В химическом отношении плазматическая мембрана представляет собойлипопротеиновый комплекс.Основными компонентами являются липиды (около 40%), белки (более 60%) и углеводы (около 2-10%).
Плазматическая мембрана выполняет ряд важнейших функций:
Барьерная. Барьерная функция плазматической мембраны заключается в ограничении свободной диффузии веществ из клетки в клетку, предотвращении утечки водорастворимого содержимого клетки. Но поскольку клетка должна получать необходимые питательные вещества, выделять конечные продукты метаболизма, регулировать внутриклеточные концентрации ионов, то в ней образовались специальные механизмы переноса веществ через клеточную мембрану.
Транспортная. К транспортной функции относится обеспечение поступления и выведения различных веществ в клетку и из клетки. Важное свойство мембраны - избирательная проницаемость, или полупроницаемость. Она легко пропускает воду и водорастворимые газы и отталкивает полярные молекулы, такие как глюкоза или аминокислоты.
Существует несколько механизмов транспорта веществ через мембрану:
пассивный транспорт;
активный транспорт;
транспорт в мембранной упаковке.
Пассивный транспорт. Диффузия - это движение частиц среды, приводящее к переносу вещества из зоны, где его концентрация высока в зону с низкой концентрацией. При диффузионном транспорте мембрана функционирует как осмотический барьер. Скорость диффузии зависит от величины молекул и их относительной растворимости в жирах. Чем меньше размеры молекул и чем более они жирорастворимы (липофильны), тем быстрее произойдет их перемещение через липидный бислой. Диффузия может быть нейтральной (перенос незаряженных молекул) и облегченной (с помощью специальных белков переносчиков). Скорость облегченной диффузии выше, чем нейтральной. Максимальной проникающей способностью обладает вода, так как ее молекулы малы и незаряжены. Диффузия воды через клеточную мембрану называется осмосом. Предполагается, что в клеточной мембране для проникновения воды и некоторых ионов существуют специальные "поры". Число их невелико, а диаметр составляет около 0,3-0,8 нм. Наиболее быстро диффундируют через мембрану легко растворимые в липидном бислое молекулы, например О, и незаряженные полярные молекулы небольшого диаметра (СО, мочевина). Перенос полярных молекул (сахаров, аминокислот), осуществляемый с помощью специальных мембранных транспортных белков называется облегченной диффузией. Такие белки обнаружены во всех типах биологических мембран, и каждый конкретный белок предназначен для переноса молекул определенного класса. Транспортные белки являются трансмембранными, их полипептидная цепь пересекает липидный бислой несколько раз, формируя в нем сквозные проходы. Это обеспечивает перенос специфических веществ через мембрану без непосредственного контакта с ней. Существует два основных класса транспортных белков: белки-переносчики (транспортеры) и каналообразующие белки (белки-каналы). Белки-переносчики переносят молекулы через мембрану, предварительно изменяя их конфигурацию. Каналообразующие белки формируют в мембране заполненные водой поры. Когда поры открыты, молекулы специфических веществ (обычно неорганические ионы подходящего размера и заряда) проходят сквозь них. Если молекула транспортируемого вещества не имеет заряда, то направление транспорта определяется градиентом концентрации. Если молекула заряжена, то на ее транспорт, кроме градиента концентрации, влияет и электрический заряд мембраны (мембранный потенциал). Внутренняя сторона плазмалеммы обычно заряжена отрицательно по отношению к наружной. Мембранный потенциал облегчает проникновение в клетку положительно заряженных ионов и препятствует прохождению ионов заряженных отрицательно.
Активный транспорт. Активным транспортом называется перенос веществ против электрохимического градиента. Он всегда осуществляется белками-транспортерами и тесно связан с источником энергии. В белках-переносчиках имеются участки связывания с транспортируемым веществом. Чем больше таких участков связывается с веществом, тем выше скорость транспорта. Селективный перенос одного вещества называется унипортом. Перенос нескольких веществ осуществляют котранспортные системы. Если перенос идет в одном направлении - это симпорт, если в противоположных – антипорт. Так, например, глюкоза из внеклеточной жидкости в клетку переносится унипортно. Перенос же глюкозы и Na+ из полости кишечника или канальцев почек соответственно в клетки кишечника или кровь осуществляется симпортно, а перенос С1~ и НСО антипортно. Предполагается, что при переносе возникают обратимые конформационные изменения в транспортере, что и позволяет премещать соединенные с ним вещества.
Примером белка-переносчика, использующего для транспорта веществ энергию выделившуюся при гидролизе АТФ, является Na+/К+- насос, обнаруженный в плазматической мембране всех клеток. Na+-K насос работает по принципу антипорта, перекачивая Na+ из клетки и К+ внутрь клетки против их электрохимических градиентов. Градиент Na+ создает осмотическое давление, поддерживает клеточный объем и обеспечивает транспорт сахаров и аминокислот. На работу этого насоса тратится треть всей энергии необходимой для жизнедеятельности клеток. При изучении механизма действия Na+/K+- насоса было установлено, что он является ферментом АТФазой и трансмембранным интегральным белком. В присутствии Na+ и АТФ под действием АТФа-зы от АТФ отделяется концевой фосфат и присоединяется к остатку аспарагиновой кислоты на молекуле АТФазы. Молекула АТФазы фосфорилируется, изменяет свою конфигурацию и Na+ выводится из клетки. Вслед за выведением Na из клетки всегда происходит транспорт К+ в клетку. Для этого от АТФазы в присутствии К отщепляется ранее присоединенный фосфат. Фермент дефосфорилируется, восстанавливает свою конфигурацию и К+ "закачивается" в клетку.
АТФаза образована двумя субъединицами, большой и малой. Большая субъединица состоит из тысячи аминокислотных остатков, пересекающих бислой несколько раз. Она обладает каталитической активностью и способна обратимо фосфорилироваться и дефосфорилироваться. Большая субъединица на цитоплазматической стороне имеет участки для связывания Na+ и АТФ, а на внешней стороне -участки для связывания К+ и уабаина. Малая субъединица является гликопротеином и функция его пока неизвестна.
Na+-K насос обладает электрогенным эффектом. Он удаляет три положительно заряженных иона Naf из клетки и вносит в нее два иона К+. В результате через мембрану течет ток, образующий электрический потенциал с отрицательным значением во внутренней части клетки по отношению к ее наружной поверхности. Na+/K+-насос регулирует клеточный объем, контролирует концентрацию веществ внутри клетки, поддерживает осмотическое давление, участвует в создании мембранного потенциала. Транспорт в мембранной упаковке. Перенос через мембрану макромолекул (белков, нуклеиновых кислот, полисахаридов, липопротеидов) и других частиц осуществляется посредством последовательного образования и слияния окруженных мембраной пузырьков (везикул). Процесс везикулярного транспорта проходит в две стадии. Вначале мембрана пузырька и плазмалемма слипаются, а затем сливаются. Для протекания 2 стадии необходимо чтобы молекулы воды были вытеснены взаимодействующими липидными бислоями, которые сближаются до расстояния 1-5 нм. Считается, что данный процесс активизируют специальные белки слияния (они выделены пока только у вирусов). Везикулярный транспорт имеет важную особенность - поглощенные или секретируемые макромолекулы, находящиеся в пузырьках, обычно не смешиваются с другими макромолекулами или органеллами клетки. Пузырьки могут сливаться со специфическими мембранами, что и обеспечивает обмен макромолекулами между внеклеточным пространством и содержимым клетки. Аналогично происходит перенос макромолекул из одного компартмента клетки в другой.
Транспорт макромолекул и частиц в клетку называется эндоцитозом. При этом транспортируемые вещества обволакиваются частью плазматической мембраны, образуется пузырек (вакуоль), который перемещается внутрь клетки. В зависимости от размера образующихся пузырьков различают два вида эндоцитоза - пиноцитоз и фагоцитоз. Пиноцитоз обеспечивает поглощение жидкости и растворенных веществ в виде небольших пузырьков (d=150 нм). Фагоцитоз - это поглощение больших частиц, микроорганизов или обломков органелл, клеток. При этом образуются крупные пузырьки, фагосомы или вакуоли (d-250 нм и более).
Рецепторная функция плазмалеммы. Это одна из главных, универсальных для всех клеток, является рецепторная функция плазмалеммы. Она определяет взаимодействие клеток друг с другом и с внешней средой.
Всё многообразие информационных межклеточных взаимодействий схематически можно представить как цепь последовательных реакций сигнал-рецептор-вторичный посредник-ответ (концепция сигнал-ответ). Передачу информации от клетки к клетке осуществляют сигнальные молекулы, которые вырабатываются в одних клетках и специфически влияют на другие, чувствительные к сигналу (клетки-мишени). Сигнальная молекула - первичный посредник связывается с находящимися на клетках-мишенях рецепторами, реагирующими только на определенные сигналы.