

7.3. Морфофизиологические и эволюционные особенности яиц хордовых
Происхождение яйцеклеток.Яйца хордовых состоят из яйцевой клетки, называемой иногда также яйцом, и яйцевых оболочек.Лицевые клеткиобразуются в женской половой железе —яичнике. Они проходят долгий путь развития, который начинается в эмбриональном и продолжается в репродуктивном периоде онтогенеза особей женского пола.
Первичные половые клеткивозникают на ранних этапах эмбриогенеза из энтодермальных клеток вегетативного полюса, как, например, у бесхвостых амфибий, или из энтодермальных клеток желточного мешка, как у всех амниот —пресмыкающихся, птиц и млекопитающих. Первичные половые клетки очень рано отличимы от других клеток по крупным размерам и прозрачной цитоплазме. Половые железы в этот момент только начинают формироваться.
Несколькими экспериментальными методами было показано, что первичные половые клетки мигрируют из места возникновения в развивающиеся гонады и заселяют их. У млекопитающих они перемешаются по дорсальной брыжейке (рис. 7.1),будучи способньми в этот период к амебоидному движению. У птиц миграция происходит пассивно по кровяному руслу. Не исключено действие химических аттрактантов, стимулирующих миграцию первичных половых клеток к гонадам. Существует мнение, что первичные половые клетки, оказавшись в каких-либо других участках зародыша, гибнут, но иногда могут перерождаться в опухоли.
Попав в гонады, первичные половые клетки начинают пролиферировать.
Они делятся митозом и называются овогониями.У большинства низших позвоночных овогонии сохраняют способность к делению на протяжении всего репродуктивного периода, так как, например, рыбы за один нерест выделяют тысячи яиц, земноводные —сотни. У высших позвоночных количество яиц, которые вызревают одномоментно, редко достигает 15,обычно их бывает меньше, иногда одно, чем можно объяснить и особенности овогенеза.
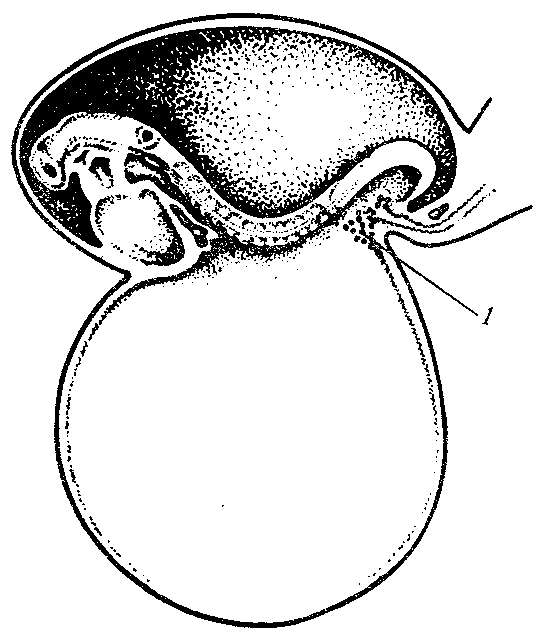
Рис. 7.1. Возникновение первичных половых клеток (I) у зародыша человека
на стадии 16сомитов
У человеческих эмбрионов женского пола овогонии размножаются наиболее интенсивно между 2-м и 5-м месяцами внутриутробного периода развития, когда их число достигает примерно 7млн. К 7-му месяцу многие овогонии погибают, а сохранившиеся входят в профазу первого деления мейоза. Для части яйцеклеток мейоз завершается в момент наступления полового созревания, а для части —непосредственно перед менопаузой.
Специфика и значение химического состава цитоплазмы яйцеклетки. Зрелая яйцеклетка, как правило, бывает крупнее овогонии и тем более крупнее любых других соматических клеток. В ходе овогенеза в цитоплазме яйцеклетки резервируется большое количество веществ, необходимых для ее созревания и обеспечения раннего эмбриогенеза. Функциональная роль запасенных веществ различна.
Во-первых,это компоненты, необходимые для процессов репликации, транскрипции и трансляции, такие, как соответствующие ферменты, рибосомы, мРНК, тРНК и их предшественники.
Во-вторых,это набор специфических регуляторных веществ, которые обеспечивают координированное функционирование всех запасенных компонентов. Некоторые из них, будучи в овоците уже к моменту оплодотворения, начинают действовать только в фазе гаструляции. К названным веществам относятся фактор дезинтеграции ядерной оболочки (с разрушения ядерной оболочки начинается прометафаза 1-го мейотического деления); фактор, вызывающий конденсацию хромосом; фактор, преобразующий ядро сперматозоида в пронуклеус и активирующий в нем синтез ДНК перед дроблением; фактор, ответственный за цитотомию во время дробления; фактор, ответственный за блок мейоза на стадии метафазы II(у многих позвоночных снятие этого блока происходит только в результате оплодотворения).
В-третьих,это желток, в состав которого входят белки, фосфолипиды, нейтральные жиры, минеральные соли. Желток представляет собой запас питательных веществ и энергетических ресурсов, необходимых для обеспечения эмбрионального периода. Многие вещества, вырабатываемые печенью, попадают в овогонии в период их роста через фолликулярные клетки яичника. Это требует от самки больших энергетических затрат.
Благодаря описанным особенностям химического состава цитоплазмы яйцеклетки зародыш на протяжении периода дробления в ряде случаев не использует для развития генетическую информацию ядер зиготы и бластомеров. Специфический химический состав и закономерное распределение веществ в цитоплазме яйцеклетки имеют большое значение для начальных фаз эмбриогенеза. Запасенные питательные и энергетические вещества обеспечивают эмбриональное развитие без дополнительного поступления их извне.
Размер яиц иего роль в эволюции. Типы яйцеклеток.В процессе развития выявляется закономерность, заключающаяся в том, что чем длиннее эмбриональный период, тем больше желтка должно быть накоплено в яйцеклетке. Продолжительность эмбрионального периода зависит от стадии, на которой зародыш переходит к самостоятельному существованию во внешней среде. Если постэмбриональное развитие идет прямым путем, т.е. без личинки и метаморфоза, то желтка в яйцеклетке должно быть больше. По количеству желтка яйцеклетки хордовых делят наалецитальные, олиго-, мезо-иполилецитальные,т.е. с ничтожно малым, малым, средним и большим количеством желтка (от греч.лецитос —желток).
У ланцетника,представителя низших хордовых, яйцеклеткаолиголецитальная.У большинства позвоночных в яйцеклетках содержится значительное количество желтка. Среди низших позвоночных (Anamnia) наиболее крупные яйца у миксин (кл. Круглоротые), у акул и химер (кл. Хрящевые рыбы) и у безногих амфибий. У осетровых рыб, а также остальныхамфибийяйцеклеткимезолецитальные,т.е. имеют среднее количество желтка. У высших позвоночных (Amniota), таких, как пресмыкающиеся, птицы и яйцекладущие млекопитающие,—полилецитальные,т.е. в яйцеклетке очень много желтка. Эмбриональное развитие у них протекает особенно долго.
Эта закономерность нарушена у сумчатых и плацентарныхмлекопитающих, которые имеютолиго-иалецитальныеяйцеклетки соответственно. У сумчатых эмбрион выходит из яйцевых оболочек и матки при незавершенном органогенезе, переносится в сумку, где и продолжает развитие. У плацентарных и человека зародыш выходит из яйцевых оболочек еще раньше, в стадии бластоцисты, но затем переходит к внутриутробному существованию, где и завершает все основные периоды развития, подготавливающие его к появлению на свет.Уменьшениеколичества желтка в яйцеклетках млекопитающих можно назватьвторичным,поскольку их предки, освоившие наземную среду, имели, как и все амниоты, полилецитальные яйца.
Полярность яйцеклеток.При малом количестве желтка в яйцеклетке он обычно распределен в цитоплазме равномерно и ядро располагается примерно в центре. Такие яйцеклетки называютизолецитальными(от греч.изос —равный). У большинства позвоночных желтка много, и он распределен в цитоплазме яйцеклетки неравномерно. Этоанизолециталъныеклетки. Основная масса желтка скапливается у одного из полюсов клетки —вегетативного полюса. Такие яйцеклетки называюттелолецйтальными(от греч.телос — конец). Противоположный полюс, к которому оттесняется свободная от желтка активная цитоплазма, называютанималъным.Если желток все же погружен в цитоплазму и не обособлен от нее в виде отдельной фракции, как у осетровых и земноводных, яйцеклетки называютумеренно телолецитальными.Если желток полностью отделен от цитоплазмы, как у амниот, то эторезко телолецитальныеяйцеклетки.
Изложенные особенности размеров и полярности яйцеклеток хордовых схематично сгруппированы в табл. 7.1.
Распределение желтка имеет решающее значение для всех последующих процессов эмбриогенеза, так как определяет будущую пространственную организацию зародыша. Вместе с тем необходимо отметить, что распределение желтка —это как бы внешнее проявление более тонких процессов поляризации яйцеклеток, осуществляющихся, по-видимому, в кортикальном слое и прилежащих к нему цитоскелетных структурах.
Таблица 7.1. Типы яйцеклеток, встречающиеся у хордовых
|
Представители типа Хордовые |
В зависимости от количества желтка |
В зависимости от распределения желтка |
|
Ланцетник Лягушка Птица Плацентарные млекопитающие (и человек) |
олиголецитальнаямезолецитальная полилецитальнаяалецитальная |
изолецитальная умеренно телолецитальная резко телолецитальная олецитальная изолецитальная |
Кортикальный слой —это поверхностный слой цитоплазмы яйца вместе с плазматической мембраной. В нем находятся микрофиламенты и кортикальные гранулы. Кортикальный слой играет важную роль в кортикальной реакции при оплодотворении.
Полярность яйцеклеток намечается еще на стадии накопления желтка в овоцитах во время их быстрого роста и закрепляется при выделении полярных телец именно на анимальном полюсе. Внутреннюю разнокачественность участков яйца называют овоплазматической сегрегацией.
Лицевые оболочки.Различаютпервичную оболочку,образуемую самой яйцевой клеткой,вторичную оболочку,являющуюся продуктом деятельности фолликулярных клеток, итретичныеоболочки,которыми яйцо окружается во время прохождения по яйцеводу.
Первичная оболочка, иногда называемая желточной, имеется у яйцеклеток всех животных. У позвоночных, в том числе млекопитающих и человека, первичная оболочка входит в состав плотной оболочки, образуя ее внутреннюю часть. Внешняя часть плотной оболочки продуцируется фолликулярными клетками и является вторичной оболочкой. Плотная оболочка изнутри пронизана микроворсинками яйцеклетки, а снаружи —микроворсинками фолликулярных клеток, отчего при большом увеличении она выглядит исчерченной и названалучистой оболочкой(coronaradiata). За свои оптические свойства у млекопитающих она получила названиеблестящей оболочки(zonapellucida). Таким образом, эта оболочка совмещает в себе первичную и вторичную оболочки.
Третичные оболочки хорошо развиты у хрящевых рыб и амфибий, но особенную сложность они приобретают у наземных позвоночных —пресмыкающихся, птиц и низших млекопитающих. Образуясь из секретов желез яйцевода, эти оболочки не имеют клеточного строения. У всех позвоночных они выполняют функции защиты зародыша от механических повреждений и действия вредных биотических факторов, таких, как бактериальные, грибковые и протозойные. Кроме того, у наземных позвоночных появляются принципиально новые функции запаса воды и питательных веществ для обеспечения нужд зародыша. У пресмыкающихся скорлуповая оболочка действует как насос, забирая воду из почвы и воздуха. У птиц запас воды находится в белковой оболочке. Поглощение и испарение воды регулируется порами в скорлуповой оболочке. Скорлупа содержит множество минеральных солей, необходимых для развития скелета зародыша.