

ТКАНИ. КЛАССИФИКАЦИЯ ТКАНЕЙ.
В основе организации высших растений лежит принцип специализации клеток, который заключается в том, что каждая клетка организма выполняет не все присущие ей функции, а только некоторые, но зато более полно и совершенно.
Ткани - устойчивые, закономерно повторяющиеся комплексы клеток, сходные по происхождению, строению и приспособленные к выполнению одной или нескольких функций.
Существуют различные классификации тканей, но все они достаточно условны.
В зависимости от основной функции различают несколько групп растительных тканей.
1. Образовательные ткани, илимеристемы, - обладают способностью к делению и формированию всех прочих тканей.
2. Покровные ткани:
• первичные;
• вторичные;
• третичные.
3. Основные ткани - составляют большую часть тела растения. Различают следующие основные ткани:
• ассимиляционные (хлорофиллоносные);
• запасающие;
• воздухоносные (аэренхима);
• водоносные.
4. Механические ткани (опорные, скелетные):
• колленхима;
• склеренхима.
5. Проводящие ткани:
• ксилема (древесина) - ткань восходящего тока;
• флоэма (луб) - ткань нисходящего тока.
6. Выделительные ткани:
• наружные:
- железистые волоски;
- гидатоды - водяные устьица;
- нектарники;
• внутренние:
- выделительные клетки с эфирными маслами, смолами, дубильными веществами;
- многоклеточные вместилища выделений, млечники.
По способности клеток к делению различают два типа тканей: образовательные, илимеристемы, ипостоянные - покровные, выделительные, основные, механические, проводящие.
Ткань называют простой, если все ее клетки одинаковы по форме и функциям (паренхима, склеренхима, колленхима).Сложные ткани состоят из клеток, неодинаковых по форме, строению и функциям, но связанных общим происхождением (например, ксилема, флоэма).
Существует также классификация тканей, основанная на их происхождении (онтогенетическая). Согласно этой классификации различают первичные и вторичные ткани. Из первичной меристемы, находящейся на верхушке побега и кончике корня, а также из зародыша семени формируются первичные постоянные ткани (эпидерма, колленхима, склеренхима, ассимиляционная ткань, эпиблема). Клетки постоянных тканей неспособны к дальнейшему делению. Из клеток специализированной меристемы -прокамбия - формируютсяпервичные проводящие ткани (первичная ксилема, первичная флоэма).
Из вторичной меристемы - камбия - формируютсявторичные ткани: вторичная ксилема, вторичная флоэма; изфеллогена образуются пробка, феллодерма, чечевички, возникающие при утолщении стебля и корня. Вторичные ткани, как правило, встречаются у голосеменных и двудольных покрытосеменных растений. Мощное развитие вторичных тканей - древесины и луба характерно для древесных растений.
ОБРАЗОВАТЕЛЬНЫЕ ТКАНИ
Образовательные ткани благодаря постоянному митотическому делению их клеток обеспечивают образование всех тканей растения, т.е. фактически формируют его тело. Любая клетка в своем развитии проходит три стадии: эмбриональную, роста и стадию дифференциации (то есть приобретения клеткой определенной функции). По мере дифференциации зародыша первичная меристема сохраняется только на верхушке будущего побега (в конусе нарастания) и на кончике корня - апикальные (верхушечные)меристемы. Зародыш любого растения состоит из клеток меристемы.
Цитологическая характеристика меристем. Типичные признаки наиболее отчетливо выражены в верхушечных меристемах. Эти меристемы составлены изодиаметрическими многогранными клетками, не разделенными межклетниками. Их оболочки тонки, содержат мало целлюлозы и способны растягиваться.
Полость каждой клетки заполнена густой цитоплазмой с относительно крупным ядром, занимающим центральное положение, и интенсивно делящейся митозом. В гиалоплазме много диффузно разбросанных рибосом, пропластид, митохондрий и диктиосом. Вакуолей немного, и они мелкие. Проводящие ткани образуются из меристемы, имеющей прозенхимную форму и крупные вакуоли, - прокамбия и камбия. Клетки прокамбия в поперечном сечении многоугольные, клетки камбия - прямоугольные.
Клетки, сохраняющие свои меристематические свойства, продолжают делиться, образуя все новые и новые клетки, называемые инициалями. Часть дочерних клеток дифференцируется, превращаясь в клетки различных тканей, их называютпроизводными инициалей. Клетки инициалей могут делиться неопределенно много раз, а производные инициалей делятся один или несколько раз и превращаются в постоянные ткани.
По происхождению различают первичные и вторичные меристемы.
Первичные меристемы
Первичные меристемы происходят непосредственно из меристемы зародыша и обладают способностью к делению. По положению в растении первичные меристемы могут быть верхушечными (апикальными), вставочными (интеркалярными) и боковыми (латеральными).
Верхушечные (апикальные) меристемы - такие меристемы, которые располагаются у взрослых растений на верхушках стеблей и кончиках корней и обеспечивают рост тела в длину. У стеблей в конусе нарастания выделяют два меристематических слоя: тунику, из которой образуются покровная ткань и периферическая часть первичной коры, и корпус, из которого образуются внутренняя часть первичной коры и центральный осевой цилиндр (рис. 2.3).
Рис. 2.3. Апикальные меристемы
стебля:а - продольный срез: 1
- конус нарастания; 2 - зачаток листа; 3 -
бугорок пазушной почки;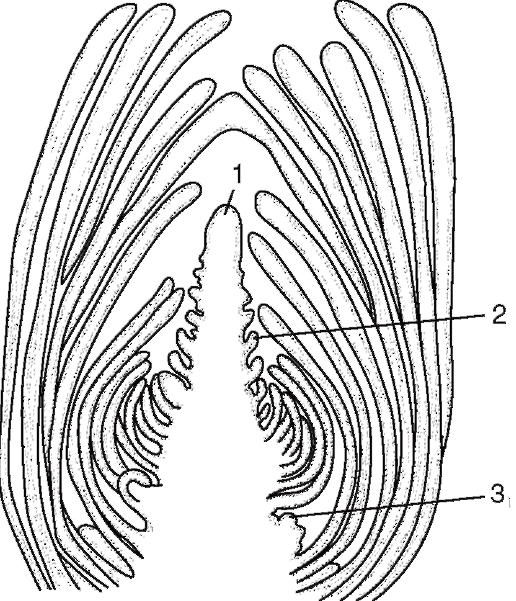
В кончике корня различают три слоя:
1) дерматоген, из которого образуется первичная покровно-всасывающая ткань - ризодерма;
2) периблему, из которой развиваются ткани первичной коры;
3) плером, образующий ткани центрального осевого цилиндра.
Боковые (латеральные) меристемы по происхождению могут быть первичными и вторичными, на поперечном срезе осевых органов имеют вид колец. Примером первичной боковой меристемы служат прокамбий и перицикл. Изпрокамбия формируются камбий и первичные элементы сосудисто-волокнистых пучков (первичная флоэма и первичная ксилема), при этом клетки прокамбия непосредственно дифференцируются в клетки первичных проводящих тканей.
Боковые меристемы располагаются параллельно поверхности органа и обеспечивают рост осевых органов в толщину.
Вставочные (интеркалярные) меристемычаще первичные и сохраняются в виде отдельных участков в зонах активного роста в различных частях растения (например, в основании черешков листьев, у оснований междоузлий). В основании междоузлий у злаков деятельность этой меристемы ведет к удлинению междоузлий, что обеспечивает рост стебля в длину.
Вторичные меристемы
К вторичным относят боковые и раневые меристемы.
Боковые (латеральные) меристемы представленыкамбием ифеллогеном. Они формируются из промеристем (прокамбия) или постоянных тканей путем их дедифференцировки. Клетки камбия делятся перегородками, параллельными поверхности органа (периклинально). Из клеток, отложенных камбием наружу, развиваются элементы вторичной флоэмы, а из клеток, отложенных внутрь, - вторичной ксилемы. Камбий, возникший из постоянных тканей путем дедифференцировки, называютдобавочньм. По строению и функции он не отличается от камбия, возникшего из промеристем. Феллоген формируется из постоянных тканей, расположенных в субэпидермальных слоях (под эпидермой). Делясь периклинально, феллоген отделяет наружу будущие клетки пробки (феллемы), а внутрь - клетки феллодермы. Таким образом, феллоген формирует вторичную покровную ткань - перидерму. Боковые меристемы располагаются параллельно поверхности органа и обеспечивают рост осевых органов в толщину.
Раневые меристемы образуются при повреждении тканей и органов. Вокруг повреждения живые клетки дедифференцируются, начинают делиться и тем самым превращаются во вторичную меристему. Их задача - образовать плотную защитную ткань, состоящую из паренхимных клеток, -каллюс. Эта ткань беловатого или желтоватого цвета, ее клетки имеют крупные ядра и достаточно толстые клеточные стенки. Каллюс возникает при прививках, обеспечивая срастание привоя с подвоем, и в основании черенков. В нем могут закладываться придаточные корни и почки, поэтому его используют для получения культуры изолированных тканей.
ПОКРОВНЫЕ ТКАНИ
Первичная покровная ткань
К первичной покровной ткани относят эпидерму собственно эпидермальных, околоустьичных, замыкающих клеток устьица и трихом.
Наружные стенки клеток эпидермы покрыты кутикулой. Она защищает клетки от механических повреждений, проникновения паразитов и поглощает ультрафиолетовые лучи. Кутинизация рассматривается как приспособление для уменьшения испарения и вымывания веществ из клеток выпадающими осадками.
Пектиновые вещества и целлюлоза, входящие в клеточную стенку, могут подвергаться ослизнению с образованием слизей икамедей. Они представляют собой полимерные углеводы, родственные пектиновым веществам, и отличаются способностью к сильному набуханию при соприкосновении с водой. Камеди в набухшем состоянии клейкие и могут вытягиваться в нити, тогда как слизи сильно расплываются и в нити не вытягиваются. Пектиновые слизи встречаются у представителей семейств лилейных, крестоцветных, мальвовых, липовых, розоцветных, в отличие от целлюлозных слизей, встречающихся гораздо реже (например, у орхидных).
Устьица представляют собой
высокоспециализированные образования
эпидермы, состоящие из двух замыкающих
клеток бобовидной формы и устьичной
щели (своеобразного межклетника между
ними). Имеются главным образом в листьях,
но встречаются и в стебле (рис. 2.6).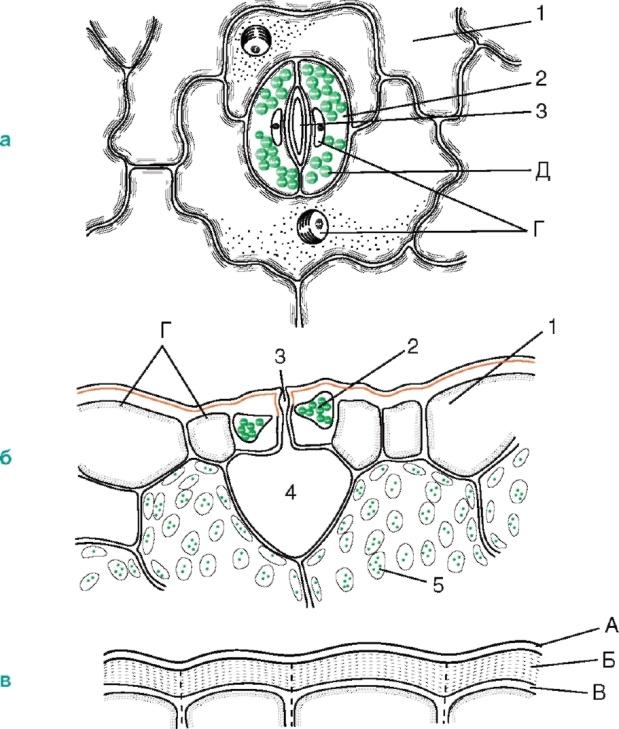
Рис. 2.6. Строение устьица:а, б - кожица листа тимьяна (вид сверху и на поперечном срезе);в - кожица со стебля цереуса (семейство кактусовых); 1 - собственно эпидермальные клетки; 2 - замыкающие клетки устьица; 3 - устьичная щель; 4 - воздухоносная полость; 5 - клетки хлорофиллоносной паренхимы; А - кутикула; Б - кутикулярный слой - оболочка с суберином и воском; В - целлюлозный слой стенки; Г - ядро с ядрышком; Д - хлоропласты
Стенки замыкающих клеток утолщены неравномерно: стенки, направленные к щели (брюшные), значительно утолщены по сравнению со стенками, направленными от щели (спинными). Щель может расширяться и сужаться, регулируя транспирацию и газообмен. Под щелью расположена крупная дыхательная полость (межклетник), окруженная клетками мезофилла листа.
Замыкающие клетки окружены околоустьичными
клетками, которые совместно
образуют устьичный комплекс (рис.
2.7). Различают следующие основные типы
устьичных комплексов: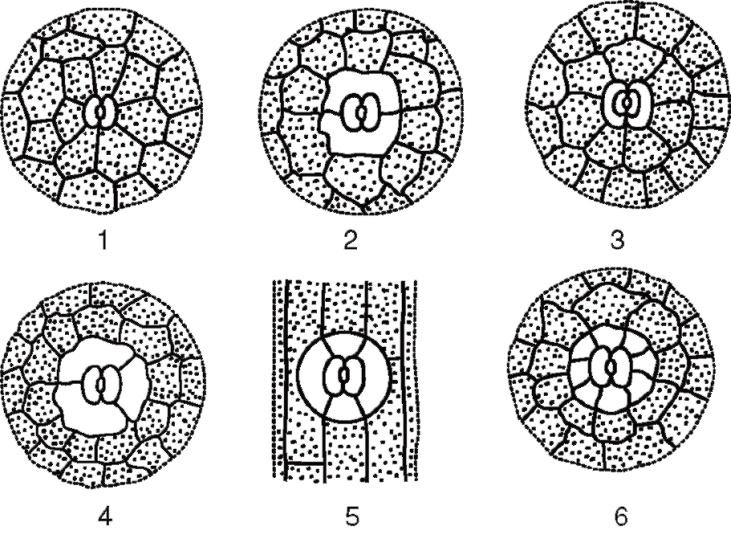
Рис. 2.7. Основные типы устьичного аппарата: 1 - аномоцитный (у всех высших растений, кроме хвощей); 2 - диацитный (у папоротников и цветковых); 3 - парацитный (у папоротников, хвощей, цветковых и гнетовых); 4 - анизоцитный (только у цветковых); 5 - тетрацитный (главным образом у однодольных); 6 - анциклоцитный (у папоротников, голосеменных и цветковых)
1) аномоцитный (беспорядочный) - замыкающие клетки не имеют ярко выраженных околоустьичных клеток; характерен для всех высших растений, исключая хвойные;
2) анизоцитный (неравноклеточный) - замыкающие клетки устьица окружены тремя околоустьичными клетками, одна из которых намного крупнее (или меньше) остальных;
3) парацитный (параллельно-клеточный) - одна околоустьичная клетка (или более) расположена параллельно замыкающим;
4) диацитный (перекрестно-клеточный) - две околоустьичные клетки расположены перпендикулярно замыкающим;
5) тетрацитный (от греч.tetra - четыре) - главным образом у однодольных;
Устьица расположены на нижней стороне
листа, но у водных растений с плавающими
листьями они находятся только на верхней
стороне листа. По форме клеток эпидермы
листа и расположению устьиц можно
отличить однодольное растение от
двудольного (рис. 2.8). Собственно
эпидермальные клетки листа двудольных
растений в очертаниях волнистые (рис.
2.9), а у однодольных они вытянутые,
ромбической формы.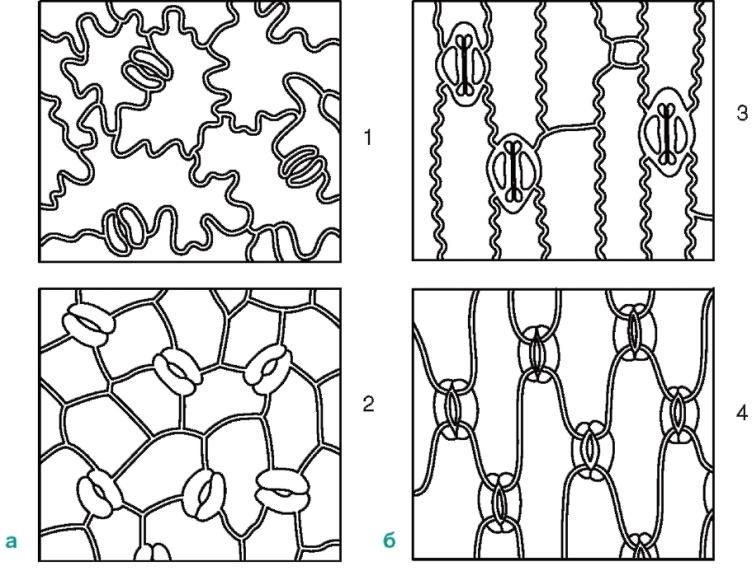
Рис. 2.8. Расположение устьиц на эпидерме (вид с поверхности):а -двудольные растения: 1 - буквица; 2 - арбуз;б -однодольные растения: 3 - кукуруза; 4 - ирис
Типы устьиц по уровню расположения относительно поверхности эпидермиса можно подразделить следующим образом.
1.7.1.Устьица, расположенные в одной плоскости с эпидермисом. Наиболее распространенный тип и обычно в описании микроскопии лекарственного растительного сырья не указывается, т.е. данный пункт опускается. Диагностическими признаками будут либо выступающие, либо погруженные устьица.
1.7.2.Выступающие устьица - устьица, расположенные над эпидермисом. Обычно при вращении микровинта микроскопа (при опускании объектива) сначала обнаруживаются такие устьица, а уже затем появляются клетки эпидермиса, поэтому на фотографии запечатлеть их с поверхности листа практически невозможно, равно как и изобразить на рисунке. В одной плоскости с эпидермисом такие устьица можно увидеть на поперечных срезах, но для этого срез должен пройти через устьице, что трудно получить при их редком расположении на листе. Такие устьица характерны, например, для листьев толокнянки.
1.7.3.Погруженные устьица - устьица, погруженные в эпидермис. При наблюдении под микроскопом при вращении микровинта (при опускании объектива) сначала четко обнаруживаются клетки эпидермиса, затем становится возможным более четко увидеть контуры устьиц. На фотографиях и рисунках препаратов с поверхности их также трудно отобразить. Встречаются в листьях ландыша, листьях вахты, листьях эвкалипта. Иногда углубления, в которых располагаются устьица, выстланы или прикрыты волосками и называютсяустьичными криптами.
1.8. Типы устьичных клеток
В литературе описывается 19 типов [1], нами выбраны только те, которые используются в анализе лекарственного растительного сырья**.
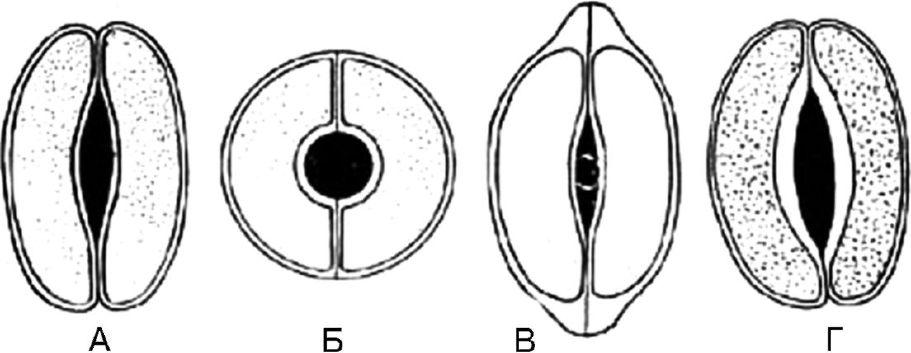
Рис. 63. Типы устьичных клеток. А - чечевицевидные; Б - сферовидные; В - колпачковидные; Г – ладьевидные
1.8.1. Чечевицевидные - 2 одинаковые клетки полулунной формы расположены симметрично. На фронтальной плоскости утолщение оболочки почти равномерное. Щель веретеновидная (рис. 63, А). Тип устьичных клеток характерен для большинства растений.
1.8.2. Сферовидные - две одинаковые, сильно кругообразноизогнутые клетки расположены симметрично. На фронтальной плоскости утолщение оболочки почти равномерное. Щель круглая (рис. 63, Б).
1.8.3.Колпачковидные - две одинаковые клетки полулунной формы в полярных частях имеют утолщения в виде колпачка. Щель веретеновидная (рис. 63, В). Встречаются у наперстянок.
1.8.4.Ладьевидные - внутренние стенки устьичных клеток утолщены. Щель веретеновидная (рис. 63, Г). Наблюдается в траве золототысячника, в листьях вахты.
Механизм работы устьиц обусловлен осмотическими свойствами клеток. При освещении поверхности листа солнцем в хлоропластах замыкающих клеток происходит активный процесс фотосинтеза. Насыщение клеток продуктами фотосинтеза, сахарами влечет за собой активное поступление в клетки ионов калия, вследствие чего концентрация клеточного сока в замыкающих клетках возрастает. Возникает разность концентрации клеточного сока околоустьичных и замыкающих клеток. В силу осмотических свойств клеток вода из околоустьичных клеток поступает в замыкающие, что ведет к увеличению объема последних и резкому возрастанию тургора. Утолщение «брюшных» стенок замыкающих клеток, обращенных к устьичной щели, обеспечивает неравномерное растяжение клеточной стенки; замыкающие клетки приобретают выраженную бобовидную форму, и устьичная щель открывается. При снижении интенсивности фотосинтеза (например, вечером) уменьшается образование сахаров в замыкающих клетках. Приток ионов калия прекращается. Концентрация клеточного сока в замыкающих клетках снижается по сравнению с околоустьичными. Вода путем осмоса уходит из замыкающих клеток, понижая их тургор, в результате ночью устьичная щель закрывается.
Клетки эпидермы плотно сомкнуты между собой, благодаря этому эпидерма выполняет целый ряд функций:
• препятствует проникновению болезнетворных организмов внутрь растения;
• защищает внутренние ткани от механических повреждений;
• регулирует газообмен и транспирацию;
• через нее выделяются вода, соли;
• может функционировать как всасывающая ткань;
принимает участие в синтезе различных веществ, восприятии раздражений и в движении листьев.
Трихомы - различные по форме, строению и функциям выросты клеток эпидермы: волоски, чешуйки, щетинки и т.п. Их подразделяют на кроющие и железистые.Железистые трихомы, в отличие от кроющих, имеют клетки, выделяющие секрет.Кроющие волоски, образуя на растении шерстистый, войлочный или иной покров, отражают часть солнечных лучей и тем самым уменьшают транспирацию. Иногда волоски находятся только там, где расположены устьица, например, на нижней стороне листа мать-и-мачехи. У некоторых растений живые волоски увеличивают общую испаряющую поверхность, что способствует ускорению транспирации.
Размеры трихом значительно варьируют. Наиболее длинные трихомы (до 5-6 см) покрывают семена хлопчатника. Кроющие трихомы имеют форму простых одно или многоклеточных, разветвленных или звездчатых волосков. Кроющие трихомы могут длительное время оставаться живыми или быстро отмирать, заполняясь воздухом.
От трихом, возникающих только при участии эпидермальных клеток, отличаются эмергенцы, в образовании которых участвуют и более глубоко расположенные ткани субэпидермальных слоев.
Анатомо-диагностические признаки, имеющие наибольшее значение и высокую вариабельность при определении лекарственного сырья. Волоски могут быть простые и головчатые, которые в свою очередь могут быть одноклеточными и многоклеточными. Многоклеточные волоски могут быть однорядными, двухрядными и ветвистыми.