

Учебник
.pdfпузыри), в среднем – криноидеи (морские лилии) и бластоидеи. По дну ползали
морские звезды и правильные древние морские ежи. Силурийский период – время расцвета криноидей.
В нижнепалеозойских породах, начиная со среднекембрийских, часто встречаются конодонты. Широкое развитие червей подтверждается наличием челюстных аппаратов – сколекодонтов и обилием ходов в глинистых породах.
Наряду с беспозвоночными в морских бассейнах ордовика и силура существовали и развивались позвоночные. В породах ордовика найдены кожные зубы бесчелюстных рыбообразных организмов. В конце силура появились первые челюстные позвоночные – настоящие рыбы.
Из низших растений широким развитием пользовались сине-зеленые водоросли, известные нам по сохранившимся в породах строматолитам и онколитам, красные, бурые и зеленые водоросли. В конце силура на суше появились высшие растения – риниофиты, подготовившие выход на сушу животных.
4.6ОРГАНИЧЕСКИЙ МИР ПОЗДНЕГО ПАЛЕОЗОЯ
Кначалу позднего палеозоя резко уменьшилось разнообразие трилобитов, граптолитов, иглокожих, табулят и наутилоидей. Появились новые группы животных и растений, обитавших не только в морях, но и на суше, определивших облик органического мира позднего палеозоя. Из беспозвоночных особенно широкого развития достигли: замковые брахиоподы, фораминиферы, четырехлучевые кораллы, гониатиты, иглокожие (криноидеи) и мшанки.
Замковые брахиоподы максимального расцвета достигли в девоне. Характеризовались богатством видов, быстрой изменчивостью во времени, что и позволило использовать их в качестве важнейшей группы для определения возраста пород. В каменноугольном периоде количество их сократилось, но все же они остались одной из доминирующих групп. По обилию продуктид, раковины которых достигали до 40 см ширины, отложения этого возраста часто называют «продуктусовыми». В пермском периоде закончили свое существование представители многих групп замковых брахиопод, поэтому граница перми и триаса легко устанавливается по этому сокращению.
Фораминиферы обитали на дне мелководных теплых морей, где образовывали массовые скопления и являлись породообразующими организмами. В девоне широкое развитие получили спирально-свернутые многокамерные формы – эндотириды, в карбоне – фузулиниды, в перми – швагериниды.
В девонском периоде дальнейшее развитие получили двузонные кораллы как одиночные, так и колониальные. В каменноугольном периоде появились
и имеет языковидную или клино- и топорообразную форму. С обеих сторон от нее свисают жабры.
На спинном (верхнем) крае обеих створок находятся макушки, с внутренней стороны которых имеются известковые отростки (зубы), разделенные межзубными ямками. В совокупности они образуют зубной (замочный) аппарат. Иногда вместо зубов присутствует связка. Замыкаются створки при помощи мускул. На внутренней стороне створок часто можно наблюдать мускульные отпечатки. Классификация двустворок осуществляется на основании строения замка.
Двустворки известны с кембрия. Наибольшее значение имеют для расчленения мезозойских и кайнозойских отложений (мел, палеоген, неоген).
Головоногие (Cephalopoda, kephale – голова, podos – нога) – высокоразви-
тые морские организмы, получившие название «приматы моря». К ним относятся: каракатицы, осьминоги, кальмары, наутилиды, вымершие аммониты и белемниты и т. д. Размеры их зачастую бывают фантастические и достигают до 6,5 м в длину и 1,95 м в поперечнике. Общее количество видов – более 10 тыс., в настоящее время обитает поряка 650 видов.
Головоногие характеризуются обособленной головой (рис. 58), высоко развитыми органами чувств и совершенными глазами. На голове моллюска располагаются многочисленные «руки» (щупальцы), служащие для захвата пищи и частично для передвижения. Число щупальцев может достигать 100. Основным средством передвижения является воронка – трубчатый мускульный орган, сообщающийся с жабрами.
а
Рис. 58. Раковина аммонита с открытым устьем
(Schindewolf, 1958);
а – аптих; в – воронка
Важной частью мягкого тела является мантия – кожистая складка, покрывающая туловище. С помощью залегающих в кайме мантии желез животное строит раковину.
182 |
171 |
Класс делится на шесть подклассов, пять из которых объединены в группу наружнораковинных, а шестой является внутреннераковинным.
Наружнораковинные имеют форму прямых, согнутых или спирально закрученных конических трубок, разделенных поперечными перегородками на камеры. Мягкое тело моллюска расположено в передней – жилой – камере. Остальные камеры заполнены газом и называются фрагмакон. Они помогают животному удержаться на воде. Во всех перегородках имеется отверстие, через которое проходит кожистая трубка – сифон. След прикрепления перегородок к стенкам раковины – перегородочная (лопастная) линия.
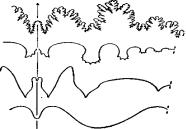
На основании строения лопастной линии, которая бывает четырех типов (рис. 59), выделяют наутилид, цератитов, гониатитов, бактритов, аммоноидей. Наутилоидеи и цератиты достигли своего расцвета в раннем палеозое, гониатиты – в позднем палеозое; мезозой – эпоха расцвета цератитов и аммонитов.
Внутреннераковинные моллюски имеют известковую или роговую внутреннюю раковину, лежащую внутри висцерального мешка. Выделяют четыре отряда: каракатицы, кальмары, осьминоги, белемниты. Белемниты, они же «громовые стрелы», «фигурные камни» – вымершие организмы, являющиеся важными руководящими формами юры и мела. В задней части тела они имели разделенную на камеры раковину – фрагмакон, к которой примыкал конический, цилиндрический или сигарообразный ростр (в народе – «чертов палец») из битуминозного радиально-лучистого кальцита. При трении ростр издавал своеобразное зловоние, напоминавшее запах кошачьей мочи, за что получил еще название «кошачий камень». Благодаря особенностям кальцитового состава, ростры очень устойчивы к выветриванию и в массовом количестве встречаются в ископаемом состоянии, образуя так называемые «поля битв белемнитов».
А) аммонитовый тип;
Б) цератитовый тип;
В) гониатитовый тип;
Г) агониатитовый тип
Рис. 59. Перегородочные линии головоногих моллюсков
Трилобиты – важнейшая в стратиграфическом отношении группа раннепалеозойских животных. Малочленистые трилобиты (миомеры) испытали расцвет в кембрии и закончили свое развитие в ордовике.
Расцвет многочленистых трилобитов (полимеров) также приходится на кембрийский период; именно в кембрийских отложениях находят остатки большинства известных родов. Начиная с позднего кембрия, полимеры постепенно теряют завоеванные ими позиции, но при этом обнаруживают прогрессивное непрерывное развитие – удивительное разнообразие их строения и внешнего облика.
Граптолиты появились в кембрии, максимального расцвета достигли в ордовике и силуре. В раннем и среднем кембрии преобладали стереостолонаты – бентосные колониальные формы, в конце кембрия появились планктонные и псевдопланктонные мономорфные колонии. В раннем ордовике получили расцвет безосые граптолиты, во второй половине ордовика – осеносные с двурядным расположением тек, в силуре – однорядные осеносные формы.
Брахиоподы обитали на мелководье. В раннем кембрии существовали исключительно беззамковые формы, в конце кембрия появились замковые брахиоподы, достигшие значительного развития в ордовике и силуре.
Головоногие моллюски – первые морские хищники – вели подвижный образ жизни. Кембрийские головоногие представлены мелкими формами, ордовикские наутилоидеи и цератитиды достигали в длину нескольких метров. Их раковина имела форму бочонка, длинной палки, рога, плоской спирали.
Кишечнополостные представлены гидроидными (строматопораты), коралловыми полипами и медузами. Строматопораты в течение ордовика и силура строили крупные колонии и наравне с другими организмами участвовали в образовании рифовых построек. Из коралловых полипов наиболее широко были распространены табуляты. Первые табуляты появились в кембрии, в ордовике широкого расцвета достигли формы с несообщающимися между собой кораллитами. В силуре к ним присоединились группы табулят с сообщающимися при помощи пор кораллитами. К концу силура табуляты достигли исключительного расцвета и получили широкое распространение во всех морях. В конце ордовика появились первые четырехлучевые кораллы. Это были простые однозонные формы. В силуре они поднялись на более высокую эволюционную ступень: развитие получили двузонные кораллы.
Важное место на морском дне занимали иглокожие. В кембрии преобладали капроидеи; в раннем ордовике к ним присоединились цистоидеи (морские
172 |
181 |
Покрытосеменные (цветковые) – наиболее высокоорганизованные растения. Они имеют семена, которые образуются внутри завязи цветка. В последствии стенки завязи превращаются в плод. К цветковым относятся пальма, дуб, береза, ясень, клен и т.д. Время расцвета – кайнозой.
Членистостебельные широко были распространены в палеозое. Они имели полый внутри членистый стебель. В карбоне и перми членистостебельные были крупнейшими древесными растениями, известными под названием каламитов. Их стволы достигали 30 м в высоту и 1 м в поперечнике. В настоящее время это исключительно травянистые растения (32 вида хвощей).
Особенно большое значение имеют семена, споры, пыльца растений, позволяющие расчленять континентальные толщи, которые практически не содержат ископаемых остатков животных. На изучении спор и пыльцы растений основан метод определения возраста континентальных отложений, получивший название спорово-пыльцевого метода.
4.5 ОРГАНИЧЕСКИЙ МИР РАННЕГО ПАЛЕОЗОЯ
Органический мир раннего палеозоя характеризуется развитием животных, низших и высших растений. Важной особенностью органического мира является способность организмов строить разные по химическому составу наружные или внутренние скелеты. Эта особенность может быть объяснена, вопервых, появлением хищников (скелет – защитная реакция организмов), вовторых, изменением химического состава вод океанов и, в-третьих, переходом от планктонного (в докембрии) образа жизни к бентосному (скелет – опора организму в зоне подвижных вод).
Особенно широко в раннепалеозойских морях были развиты археоциаты, трилобиты, граптолиты, головоногие моллюски, коралловые полипы и иглокожие. Возможно, в водах обитали и другие беспозвоночные, но у одних групп остатки не сохранились (черви), а другие только начали свое историческое развитие и не сыграли существенной роли. Из этих групп можно назвать двустворок, гастропод, остракод, тентакулитов, мшанок, ракоскорпионов и водных скорпионов. Общий специфический облик раннепалеозойского органического мира морских беспозвоночных дополняли появившиеся еще в протерозое простейшие – фораминиферы, радиолярии, губки.
Археоциаты населяли теплые мелководные моря. Часто они сплошным ковром покрывали дно и принимали участие в построении рифов. Максимального расцвета достигли в раннем кембрии, к концу кембрия вымерли.
Тип членистоногие (Arthropoda). Данный тип объединяет примерно ¾ видов всего животного царства. Это насекомые, раки, пауки, крабы, скорпионы, клещи, вымершие трилобиты и т. д. – организмы, тело которых состоит из большого количества члеников или сегментов. Тип разделен на 4 подтипа: трилобитообразные, жабродышащие, хелицеровые и трахейные.
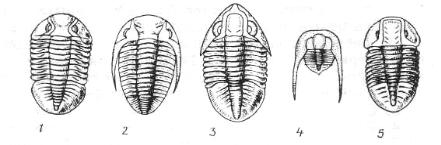
В подтипе трилобитообразных наиболее изучены трилобиты – вымершие животные, обитавшие на морском дне. Массовое распространение они имели в раннем палеозое. Хитиново-известковый панцирь трилобитов разделен на 3 части (рис. 60): головной щит, туловищный отдел и хвостовой щит. Туловищный отдел состоит из сегментов, число которых у разных видов изменяется от 2 до 44. Каждый сегмент снабжен парой конечностей. Конечности (4-5 пар) располагаются и на нижней стороне головного щита. Трилобиты с количеством сегментов 2-4 имеют размеры 1-2 см и относятся к подклассу малочленистых (Miomera). К многочленистым (Polimera) относятся трилобиты с числом сегментов 5-44, их длина может достигать 75 см. Руководящие формы кембрия, ордовика, силура.
а 
б
в
Рис. 60. Руководящие формы трилобитов песчано-известковых фаций мелководья (по Г. Крумбигель и Х. Вальтер):
1 – Asaphus expansus; 2 – Chasmops odin.; 3 – Megistaspis limbata; 4 – Cryptolithus goldfussi; 5 – Niobe sp.;
а– головной щит; б – туловищный отдел; в – хвостовой отдел
Кподтипу жабродышащих относят класс ракообразных (Crustacea), из которых наибольшее значение имеют отракоды (Ostracoda, рис. 61) – ракушковые рачки. Их тело заключено в раковину, поверхность которой либо гладкая, либо скульптированная, покрытая бугорками, шипами, ребрами. На голове располагаются 3 глаза, 2 пары конечностей, антенны и антеннулы. Тело несет от 1 до 3 пар конечностей.
180 |
173 |
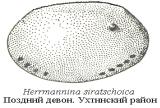
Остракоды – часто встречающиеся и морфологически разнообразные фоссилии (рис. 61). Многие из них являются руководящими формами; в силуре и девоне были породообразующими организмами. В нефтяной отрасли они используются для расчленения и корреляции осадочных толщ, служат хорошими индикаторами условий осадконакопления.
Рис. 61. Род Herrmannina siratschoica (увел.). Остракода из сирачойской свиты верхнего девона, г. Сирачай, Ухтинский  район, Южный Тиман
район, Южный Тиман
Подтип хелицеровые – это преимущественно наземные животные (клещи, скорпионы, пауки) и редкие морские формы (мечехвосты). Тело отчетливо дифференцируется на головогрудь и брюшко. В ископаемом состоянии известны с кембрия.
Подтип трахейные объединяет наземных насекомых и многоножек, дышащих при помощи трахеи. Появились в девоне, максимального расцвета достигли в кайнозое. Стратиграфического и породообразующего значения не имеют.
Тип мшанки (Bryozoa). Морские прикрепленные колониальные животные, покрывающие подводные предметы в виде корочки, пленки, мха или лишайника. Представлены более чем 900 родами, разделенными на 2 класса: голоротые и покрыторотые. Мшанки появились в конце кембрийского периода. Начиная с ордовика, а особенно в поздней перми, мелу и в настоящее время они являются важнейшими рифостроителями. Стратиграфическое значение ограничено.
Тип плеченогие (Brachiopoda). Брахиоподы – одиночные морские живот-
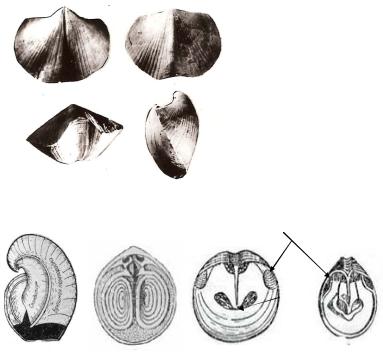
ные, прикрепленные ко дну или к какому-нибудь предмету с помощью ноги. Тело заключено в неравностворчатую раковину (рис. 62): брюшная створка выпуклая, более крупная, чем спинная. Внутренняя поверхность раковины выстлана мягкой перепонкой – мантией. Большая часть тела расположена в задней примакушечной части. В передней части раковины находятся ротовое отверстие и пара спиральных тяжей в виде петли или спирали, покрытых ресницами. Это ручной аппарат, предназначенный для сбора пищи и частично выполнящий функции дыхания. Створки раковины сочленяются при помощи специальных мускулов или замка, который состоит из двух зубов, расположенных на внутренней стороне брюшной створки, и двух ямок под макушкой спинной створки (рис. 63).
Низшие растения состоят из клеток. К ним относятся бактерии, водоросли, кокколитофориды, грибы. Тело их не подразделяется на ткани и представляет собой так называемое слоевище. Наибольшее значение имеют водоросли (сине-зеленые, диатомовые, бурые) и кокколитофориды – водные организмы, клетки которых способны выделять кальцит или кремнезем, за счет которых формируются такие горные породы, как известняки, трепел, опоки, писчий мел. Известны они с кембрия.
Высшие растения впервые появились на суше на границе силура и девона. В их теле выделяются стебель, листья и корень. Высшие растения подразделяют на 5 типов: мохо-, псилофито-, плауно-, папоротниковидные и членистостебельные.
Моховидные впервые появились в карбоне, максимального расцвета достигли в конце кайнозоя.
Псилофитовидные – низкоорганизованные растения, среди которых были и чисто водные растения и частично амфибиальные. Воздушные побеги отдельных видов псилофитов могли достигать в длину 1 м. Появились в конце силура, к концу девона вымерли.
Тип плауновидные – обширная группа споровых растений, большинство из которых представлено древовидными формами. Максимального расцвета достигли в каменноугольный период. Некоторые из них, например, лепидодендроны и сигиллярии, достигали высоты 30-40 м при диаметре ствола в 2 м, тогда как современные их виды представлены исключительно мелкорослыми травянистыми растениями.
Папоротниковидные, наиболее многочисленный и разнообразный тип, подразделяется на безсеменные (папоротники), голо- и покрытосеменные. Безсеменые размножаются при помощи спор. Наименее развитые формы имели место в девонском периоде. В карбоне им на смену пришли довольно крупные растения с богатой кроной. В современной флоре крупные папоротники встречаются только в тропическом поясе.
К голосеменным относятся растения, размножающиеся семенами, которые образуются либо в пазухах листьев, либо в шишках. Это семенные папоротники и их разновидность птеридоспермиды, кордаиты (мощные деревья с лентообразными листьями), гинкговые, цикадовые, хвойные. Семенные папоротники и кордаиты были распространены в палеозое, гинкговые и цикадовые – в мезозое. Первые хвойные появились в ранней перми. Ель, сосна, кедр и т.п. широко развиты и в современной флоре.
174 |
179 |
В ископаемом состоянии чаще всего встречаются граптолиты («письменные камни») – морские колониальные животные, представляющие собой систему расчлененных на камеры трубочек. Колонии прямые, изогнутые, спиралевидные, сетчатые, древовидные. Появились в кембрии, максимального расцвета достигли в ордовике и силуре, вымерли в каменноугольный период.
Тип конодонты (Conodonta). Это животные длиной до 40 мм, шириной до 2 мм, не имевшие скелетных образований, кроме зубовидных (конодонт). Жили со среднего кембрия до триаса. Используются для расчленения отложений палеозоя.
Тип хордовые (Сhordata). Хордовые – двухсторонне-симметричные высокоорганизованные животные, внутри тела которых имеется внутренний осевой скелет. У примитивных хордовых – это упругий хрящевой стержень (хорда), у высокоорганизованных – позвоночный столб, состоящий из серии хрящевых и костных позвонков, снабженных верхними и нижними костными дугами. Первые защищают спинной мозг, вторые – внутренние органы. На теле имеется покров (кожный, чешуйчатый, волосяной), который предохраняет организмы от излишнего испарения, механических повреждений, проникновения бактерий и пр.
Тип хордовых делят на 3 подтипа: оболочники, бесчерепные и позвоночные. Подтип позвоночные объединяет бесчелюстных (панцирные или щитковые животные, похожие на рыб, но не имеющие челюстей и парных плавников), рыб, земноводных (амфибии), пресмыкающихся (рептилии), птиц и млекопитающих. Позвоночные имеют сложно устроенные органы пищеварения, дыхания, кровеносную и нервную системы.
Изучение ископаемых остатков хордовых дает возможность проследить эволюцию высших форм, в том числе человека. Однако большого стратиграфического значения они не имеют, что объясняется условиями обитания. Большая часть хордовых, за исключением бесчелюстных, рыб, некоторых земноводных и пресмыкающихся (ихтиозавры, мезозавры, плезиозавры) обитают на суше, где условия захоронения остатков менее благоприятны, чем в море.
4.4.2 Царство растений
Развитие растительного царства всегда опережало развитие животного мира. Это может быть объяснено тем, что жизненная устойчивость растений создает основу для развития животных. Первые следы жизни растений известны из пород с возрастом 3,5 млрд лет.
Внутренне строение плеченогих сложное: развиты нервная, кровеносная, пищеварительная и половая системы. В зависимости от внутреннего строения и формы раковины их делят на 2 класса: беззамковые и замковые. Всего в природе существует порядка 950 отрядов.
|
|
|
|
|
б |
|
а |
|
|
|
Рис. 62. Внешний вид раковин |
||
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
брахиопод. Род Uchtospirifer |
|
|
|
|
|
|
timanicus, тиманская свита, |
|
|
|
|
|
|
Южный Тиман, Ухтинский район: |
|
|
|
|
|
|
а – брюшная створка с глубоким |
|
|
|
|
г |
|
синусом; б – спинная створка с |
в |
|
|
|
|
||
|
|
|
|
|
угловато-округленным седлом; |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
в – вид со стороны переднего |
|
|
|
|
|
|
края; г – вид сбоку |
а |
|
|
|
г |
|
б |
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
д
в
Рис. 63. Внутреннее строение раковин брахиопод:
а – схема внутренних пластинок; б – схема спиралевидного «ручного» аппарата; в – внутренняя сторона брюшных створок; г – отпечатки мускулов; д – отпечатки «ручного» аппарата
Геологическое значение брахиопод велико. Многие их представители занимали ведущее место среди морских животных палеозоя и дали много руководящих форм. Стратиграфия палеозоя в значительной степени базируется на изучении остатков плеченогих. Кроме того, они играли и играют важную роль в породообразовании.
178 |
175 |

Тип иглокожие (Echinodermata). К иглокожим относятся морские одиночные животные, имеющие радиальную пятилучевую или двустороннюю симметрию мягкого тела. Они имеют внутренний известковый, построенный из пластинок скелет, несущий иглы. Это высокоорганизованные организмы с пищеварительной, кровеносной и нервной системами. Специфичной для этого типа является амбулакральная (водно-сосудистая) система, служащая для обеспечения тока воды, дыхания и притока пищи. Известны с кембрия. В настоящее время насчитывается до 500 видов; около 13 видов вымерло. Скопления остатков иглокожих могут быть столь велики, что образуют эхинодермовую брекчию, эхиносферитовые и криноидные известняки.
Тип делят на 2 подтипа: прикрепленные и свободноживущие. Прикрепленные (стебельчатые) – морские пузыри – цистоидеи (Cystoidea,
рис. 64), морские лилии – криноидеи (Crinoidea, рис. 65), морские бутоны – бластоидеи (Blastoidea) ведут прикрепленный донный образ жизни, реже свободно лежат на дне. Морские лилии напоминают растения, скелет которых состоит из большого числа члеников и пластинок.
Рис. 64. Морские пузыри – цистоидеи, обитатели морей ордовикского периода: 1 – представитель рода Homocystistes; 2 – представитель рода Pleurocystistes;
а – «руки», б – чашечка, в – стебель
Рис. 65. Палеозойские морские лилии из девонских отложений Эйфеля:
1 – представитель рода Codiacrinus;
2 – представитель рода Cupressocrinus
Морские пузыри имели шарообразный или грушевидный скелет, выполненный из многоугольных известковых пластинок. Морские бутоны обладали пятилучевой симметрией чашечки, имеющей форму бутона и короткий стебель. Почти все прикрепленные иглокожие – древние животные: жили и вымерли в палеозое, лишь некоторые виды морских лилий обитают в современных морских бассейнах.
Свободноживущие иглокожие ведут подвижный бентосный образ жизни. Передвигаются по морскому дну при помощи амбулакральных игл и ножек. К ним относятся морские ежи, морские звезды, морские огурцы (голотурии).
Морские ежи, в народе известные как «ведьмины камни» – подвижные организмы шарообразной, дисковидной, овальной или сердцевидной формы с таблитчатым скелетом, сверху покрытым кожей. Таблички несут подвижные иглы, отпадающие после смерти, и поры для выхода ножек. Ротовое отверстие расположено на выположенной стороне панциря центрально или эксцентрично. Анальное отверстие лежит либо на выпуклой стороне, на ее макушке, либо смещено назад и вниз. По особенностям положения рта и анального отверстия морских ежей делят на правильные (рот – в центре нижней стороны, анальное отверстие – в центре сверху) и неправильные (рот и анальное отверстие смещены). Руководящие формы мела.
Морские звезды – донные активно подвижные формы с плоским звезднообразным строением. Их руки, служащие для передвижения, радиально отходят от центрального диска. В ископаемом состоянии встречаются редко, преобладают современные виды.
Тип полухордовые (Hemichordata). Объединяет 3 класса: кишечнодышащие, крыложаберные, граптолиты (рис. 66).
ветвь (рабдосома)
|
|
|
|
ось |
|
|
Рис. 66. Схематическое изображение |
|
|
|
|
(виргула) |
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
однорядного граптолита |
|
теки( ) |
|
|
|
|
|
|
|
|
|
|
|
|
|
(по А. Г. Мюллеру, увел.) |
|
|
|
|
|
|
|
|
|
камеры |
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
эмбриональная камера |
|
||
|
|
|
|
(сикула) |
|
||
|
|
|
игла (виргелла) |
|
|
||
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
176 |
177 |
Условные обозначения к рис. 76:
КСТМ – Тиманский сложный мегавал; ВВМ – Верхневымская депрессия; ВТМ– Восточно-Тиманскиймегавал; СПМ– Северо-Печорскаямоноклиналь; МКМ – Малоземельско-Колгуевская моноклиналь; НС – Нерицкая ступень; ЕСС – Ерсинская синклинальная структура; ИС – Ижемская ступень; ОЛС – Омра-Лыжская седловина; ПКМ – Печоро-Кожвинский мегавал; ДП – Денисовский прогиб; КМ – Колвинский мегавал;ХВ – Хорейверская впадина; АСЗ – Варандей-Адзьвинская структурная зона; Предуральский прогиб: ВПП – Воркутинское поперечное поднятие; ГЧ– грядаЧернышева; КРВ– Косью-Роговскаявпадина; БС– Большесынинская впадина; СПП– Среднепечорскоепоперечноеподнятие; ВПВ– Верхнепечорская впадина; ППП– Полюдовскоепоперечноеподнятие; СВ– Соликамскаявпадина
Ордовикская система развита только на территории Ленинградской, Новгородской и Псковской областей и залегает на размытой поверхности кембрийской. Представлена песчаниками и известняками с прослоями диктионемовых (с отпечатками граптолитов) и горючих сланцев. Мощность 100-350 м. На территории Печорской плиты с красноцветным терригенно-карбонатным вендскоордовикским комплексом (мощность до 1,0 км) связаны проявления и месторождения нефти и газа.
В силуре платформа испытала всеобщее поднятие, мелкое море заходило лишь на территорию Северного Тимана и Печорской впадины. На большей части платформы наступил длительный континентальный перерыв, продолжавшийся и в течение раннедевонской эпохи. На Северном Тимане и полуострове Канин силурийские отложения залегают на метаморфических сланцах рифейского фундамента и сложены песчаниками, сланцами, известняками и доломитами. На Северном Тимане в основании силура залегает пачка базальных конгломератов с галькой кварца и обломками рифейских сланцев. Максимальная мощность (п-ов Канин) 180 м. В Ижма-Печорской синеклизе и ПечороКолвинском авлакогене разрезы силура преимущественно карбонатные.
Нижнедевонские красноцветные песчано-глинистые отложения континентального происхождения установлены в глубоких впадинах ВолгоУральской антеклизы. В пределах Тиманской гряды к нижнему девону относят косо-и горизонтально слоистые кварцевые песчаники с линзами и прослоями алевролитов, гравелитов и конгломератов, сохранившие отпечатки мягкотелых животных, чешую рыб, растительный детрит. Мощность 150 м. В нижнедевонских терригенно-карбонатных толщах Печоро-Колвинского авлакогена зафиксированы нефтепрявления содержат промышленные скопления углеводородов (Харьягинский и Возейский валы).
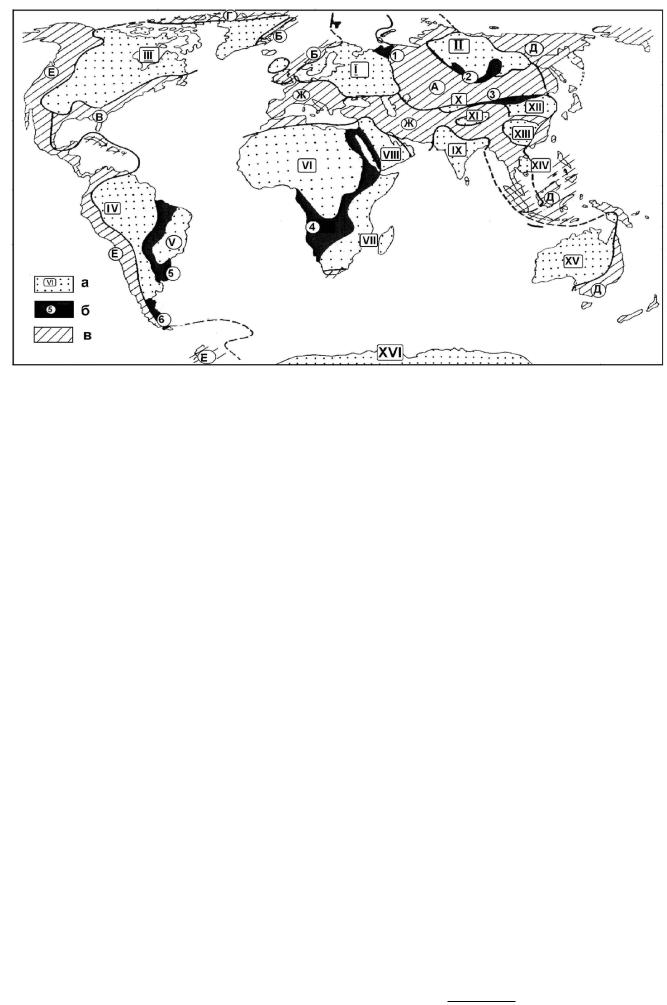
отдельные глыбы (срединные массивы). К началу позднего протерозоя в структуре литосферы существовали следующие эпикарельские платформы (рис. 67): Восточно-Европейская (1), Сибирская (II), Северо-Американская (III), ЮжноАмериканская (IV), Восточно-Бразильская (V), Северо-Африканская (VI), Юж- но-Африканская (VII), Аравийская (VIII), Индостанская (IX), Таримская (X), Тибетская (XI), Китайско-Корейская (XII), Южно-Китайская (XIII), Индосинийская (XIV), Австралийская (XIV), Антарктическая (XV).
Породы, сформировавшиеся на этом этапе, принимают участие в строении всех древних платформ вместе с архейскими породами. Среди них выделяют два существенно различных комплекса:
1)геосинклинальный, сложенный метаморфизованными осадочными и вулканическими породами с интрузиями. Заполняет широкие полосы
изоны, разделенные архейскими массивами;
2)протоплатформенный, сложенный осадочно-вулканогенными толщами. Геосинклинальные нижнепротерозойские складчатые комплексы зани-
мают обширные пространства на Восточно-Европейской платформе, в частности: на Балтийском щите, на юго-западе Северо-Африканской платформы, на Аравийском полуострове. Узкие складчатые системы и зоны располагаются на Канадском щите, на Южно-Американской платформе. Это: гнейсы, образовавшиеся при метаморфизме песчано-глинистых толщ, кварциты, филлиты, карбонатные образования, кислые и средние вулканиты, толщи метаморфизованных основных эффузивов. Породы дислоцированы, прорваны многочисленными гранитоидными интрузиями. Мощность геосинклинальных отложений 8-10 км.
Отложения протоплатформенного чехла покрывают ограниченные участки всех платформ, достигая при этом огромных мощностей (до 10 км). Они представлены продуктами размыва архейского кристаллического основания – песчаниками с прослоями и пачками конгломератов, глинистыми и карбонатными породами. Их сопровождают туфовые и лавовые серии различного состава, а также интрузии габбро, норитов, крупные тела гранитоидов. Породы сильно деформированы, образуют складчатые структуры, системы приразломных складок, ограниченные разломами блоки, моноклинали и т. п.
К началу позднего протерозоя сформировалась карельская структура земной коры. Карельские образования – карелиды явились ядрами будущих континентов.
224 |
193 |
194
Рис. 67. Схема строения Земли к началу палеозоя: а – эпикарельские платформы (см. гл.4.9.1.3, с. 196);
б– области байкальской складчатости (гл. 4.9.1.4, с. 199); в – геосинклинальные пояса и геосинклинали:
А– Урало-Монгольский; Б, В – Атлантический (Б – Грампианская, В – Аппалпчинская геосинклинали);
Г– Арктический (Иннуитская геосинклиналь); Д, Е – Тихоокеанский; Ж - Средиземноморский
223 |
структуры Тектонические .76 .Рис |
|
плиты Печорской |
Ижма-Печорская синеклиза занимает центральную часть плиты и осложнена Малоземельско-Колгуевской моноклиналью, Нерицкой ступенью, Ерсинской синклинальной структурой, Ижемской ступенью, Омра-Лыжской седловиной.
Печоро-Колвинский авлакоген расположен на запади плиты. В его пределах выделены следующие структуры: Северо-Печорская моноклиналь; ПечороКожвинский мегавал, Денисовский прогиб, Колвинский мегавал, Хорейверская впадина, Варандей-Адзьвинская структурная зона.
5.2.1.3 Стратиграфия осадочного чехла платформы
Формирование типичного платформенного чехла началось в рифее. Рифейские отложения присутствуют в окраинных депрессиях и глубоких авлакогенах, представлены красноцветными и пестроокрашенными терригеннокарбонатными толщами мощностью до 3,5 км и более. Формирование авлакогенов сопровождалось магматизмом траппового типа. На кристаллическом основании платформенные образования залегают резко несогласно и, в отличие от осадков фундамента, не дислоцированы. Органические остатки (строматолиты) встречаются крайне редко.
Впервую половину венда структурный план платформы не изменился. Разнозернистые терригенные и карбонатные осадки заполняли авлакогены. Во второй половине венда узкие прогибы переросли в пологие впадины. Почти вся платформа стала ареной осадконакопления, за исключением северной части Балтийского щита. Осадконакопление сопровождалось излияниями базальтовой лавы. Мощность отложений венда, представленных конгломератами, гравелитами, песчаниками, алевролитами и аргиллитами с пачками карбонатных пород, 400-500 м.
ВТимано-Печорской области с вендом связано образование молассоидных терригенных и грубообломочных вулкано-терригенных толщ. Накопившиеся породы смяты в складки, разбиты системой глубинных разломов, в отдельных местах прорваны гранитными интрузиями.
Скембрия до среднего девона на территории платформы господствовал континентальный режим. В кембрии и ордовике мелководный морской бассейн существовал только в Балтийском прогибе, занимавшем территорию от Баренцева моря до широты г. Котласа, и в пределах Печорской низменности. Кембрийские отложения характеризуются преимущественно песчано-глинистым составом и имеют мощность от 100 до 250 м.
4.9.1.4 Позднепротерозойский этап развития Земли
Позднепротерозойский этап (1,6-0,542 млрд лет) – это этап совместного существования в литосфере океанических и континентальных структур, геосинклиналей и платформ. С этого момента и на протяжении всей последующей истории одновременно существуют участки типично платформенного и типично геосинклинального режима, т. е. земная кора вступает в платформен- но-геосинклинальную стадию своего развития, начинают оформляться основные черты ее современного структурного плана.
В рифее режим древних платформ стабилизировался, в их пределах затухли интенсивная тектоническая деятельность, процессы гранитизации и метаморфизма. Платформы испытывали, только вертикальные движения. Происходило дальнейшее дробление их по разломам на блоки. Заложились многочисленные авлакогены, которые заполнялись красноцветными континентальными песчаниками, гравелитами и глинами с прослоями эффузивов. Этот период становления древних платформ выделяют как авлакогеновую стадию их развития. В венде отложения чехла вышли за границы авлакогенов, образовав овальные депрессии – синеклизы. Авлакогеновая стадия развития древниех платформ сменилась стадией синеклиз. Древние геосинклинальные системы, сохранившиеся от архея и раннего протерозоя, продолжалиунаследованноразвиваться.
Поздний протерозой ознаменовался рядом эпох складчатости: это готская и эльсонская, проявившиеся в Европе и Северной Америке, дальсландская, кибарская и минасская – в Европе, Африке и Южной Америке и заключительная – байкальская (700–500 млнлет). Байкальская складчатость явилась чрезвычайно важной вехой в истории развития Земли. Ее проявление привело к окончательному оформлению древних платформ в известных в настоящее время границах. В конце протерозоя к древним платформам присоединились эпибайкальская Печорская плита (1) и эпибайкальские складчатые области: Енисейско-Байкальская (2), Внутримонгольская (3), Внутриафриканская (4), Внутрибразильская (5), Патагонийская (6, рис. 67). Вместе они образовали докембрийские платформы – участки земной коры с гетерогенным по возрасту фундаментом: карельским на основной территории и байкальским в пределах окраин (рис. 68). На протяжении всей последующей историиониразвивалиськакединоецелое.
Платформы южного полушария объединились и сформировали единый крупный массив, названный Гондвана. Пространство между платформами занимали Средиземноморский подвижный пояс (океан Прототетис), СевероАтлантический (океан Япетус), Урало-Монгольский (Палеоазиатский океан), Тихоокеанский (океан Протопасифик) и Арктический подвижные пояса.
222 |
195 |
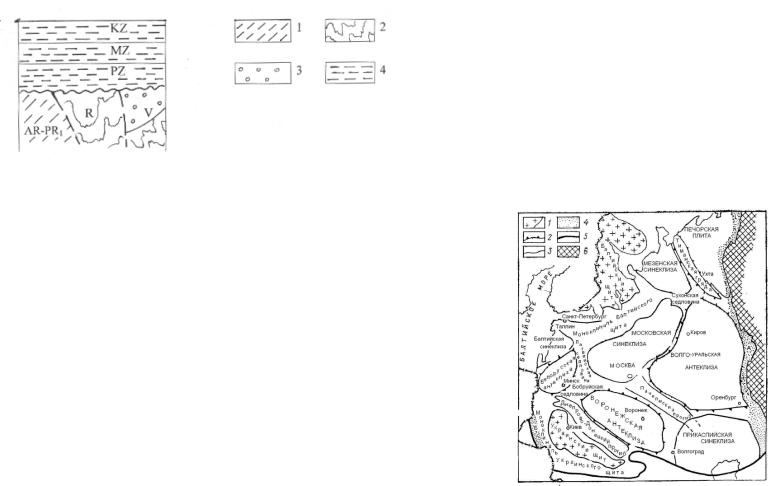
Рис. 68. Схема строения докембрийской платформы: структурно-формационные комплексы: 1 – основание геосинклинального комплекса; 2 – собственно геосинклинальный комплекс; 3 – орогенный комплекс; 4 – платформенный чехол
В пределах подвижных поясов продолжались активные тектонические, магматические и метаморфические процессы. На протяжении всей последующей истории заложение геосинклинальных систем происходило только в пределах этих поясов. Основная закономерность их развития – сужение площадей с геосинклинальным режимом и расширение областей с платформенным режимом за счет замыкания отдельных систем, преимущественно периферических, и образование на их месте молодых платформ.
Древние платформы играли в этом процессе роль ядер консолидации. К началу палеозоя сформировалась байкальская структура земной коры, которая включала древние платформы; байкалиды, геосинклинальные пояса.
Поздний протерозой – это начало формирования на платформах типичного платформенного чехла, в котором в последующем определились два комплекса: рифейский и вендский. Рифейский комплекс не имеет сплошного распространения и распространен только в авлакогенах, вендский – входит в состав сплошного чехла.
На территории подвижных поясов среди массивов ранней консолидации шло развитие многочисленных геосинклинальных систем, обладавших всеми чертами типичных геосинклиналей. Обособились эвгеосинклинальные и миогеосинклинальные прогибы с образованием формаций соответствующего ряда.
Физико-географические условия протерозоя существенно отличались от тех, которые установились в архее. В атмосфере происходило дальнейшее увеличение содержания свободного кислорода и сокращение содержания углекислоты. По дан-
ленную от Балтийского щита Балтийской моноклиналью. Восточная часть плиты характеризуется более глубоким залеганием фундамента и наличием мощного осадочного чехла. Здесь выделяются две синеклизы – Московская, простирающаяся на северо-востоке почти до Тимана, и юго-восточная Прикаспийская. Их разделяет сложно построенная погребенная Волго-Уральская антеклиза. Ее фундамент расчленен на выступы (Токмовский, Татарский и др.), между которыми располагаются грабены-авлакогены (Казанско-Сергиевский, Верхнекамский и др.) С запада Волго-Уральская антеклиза обрамлена Сухонской седловиной. Между Волго-Уральской и Воронежской антеклизами простирается глубокий Пачелмский рифейский авлакоген, сливающийся на севере с Московской синеклизой (рис. 75). Здесь фундамент плиты погружен на глубину 3-5 км, в Прикаспийской впадине – на 20-25 км.
Рис. 75. Схема региональной тектоники ВосточноЕвропейской платформы:
1 – щиты; 2-3 – границы: 2 – региональных поднятий, 3 – синеклиз; 4 – передовые прогибы:
а – Предуральский, б – Предкарпатский;
5 – южнаяграницаплатформы;
6 – Уральская горная страна
На крайнем севере Русской плиты между Балтийским щитом и Тиманской грядой фиксируется Мезенская синеклиза.
В пределах Печорской эпибайкальской плиты выделяют: Тиманскую гряду, Ижма-Печорскую синеклизу, Печоро-Колвинский авлакоген (рис. 76).
Тиманская гряда (кряж) – крупная структура, являющаяся западным ограничением Тимано-Печорской нефтегазоносной провинции и Ижма-Печорской синеклизы. В строении Тиманской гряды выделяют Канино-Северо-Тиманский сложный мегавал (КСТМ), Верхневымскую депрессию (ВВМ), ВосточноТиманский мегавал (ВТМ), осложненные поднятиями, валами, депрессиями.
196 |
221 |