

Fiziologia_rasteny
.pdfÐèñ. 2.4. Строение молекулы хлорофилла
Молекулу хлорофилла делят на две части: порфириновое ядро и фитольный хвост. Фитольный хвост в два раза длиннее ядра. Порфириновое ядро благодаря наличию атомов кислорода и азота гидрофильно. Фитольный хвост — это углеводородная, а это зна- чит — гидрофобная часть молекулы хлорофилла. Поэтому порфириновое ядро размещается в гидрофильной части мембраны тилакоида, а фитольный хвост в гидрофобной. Имея разные свойства, две части молекулы хлорофилла выполняют различную функцию: порфириновое ядро, содержащее конъюгированные связи, поглощает свет, а фитольный хвост играет роль якоря, который удерживает молекулу хлорофилла в определенной части мембраны тилакоида.
Таким образом, видим, что подобно тому, как отдельные органы многоклеточного организма или органеллы в клетке имеют определенное строение и выполняют в связи с этим особую функцию, так и особые части молекулы хлорофилла владеют специальными свойствами, а соответственно и функциями.
82
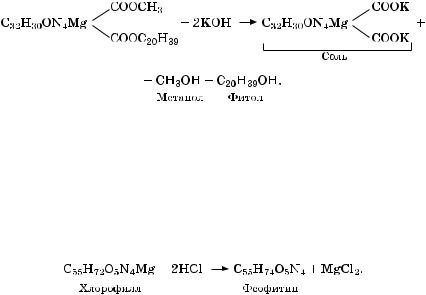
Доказать, что свет поглощается порфириновым ядром молекулы хлорофилла, можно с помощью реакции хлорофилла со щело- чью, в результате которой образуются два спирта (метанол и фитол) и соль хлорофилла:
Щелочь отсекает от молекулы хлорофилла фитольный хвост, в результате образованная соль теряет способность растворяться в бензине, но сохраняет зеленый цвет хлорофилла. Следовательно, растворимость пигмента в бензине, его гидрофобность обусловлена остатком фитола, а поглощение света связано с порфириновым ядром. Атом магния также влияет на поглощение света молекулой хлорофилла. Если с помощью кислоты заместить магний на водород, то образовавшееся вещество (феофитин) принимает крас- но-бурый цвет вместо зеленого:
Âприродных условиях образование феофитина происходит осенью, при старении листьев, под воздействием неблагоприятных факторов. В результате листья желтеют. Под влиянием различных факторов увеличивается проницаемость мембран и кислый вакуолярный сок, проникая в хлоропласты, преобразует хлорофилл в феофитин. Поскольку избирательная проницаемость мембран нарушается под воздействием разных факторов, то листья желтеют под воздействием низких и высоких температур, при дефиците воды и ее избытке и т. д.
Âнастоящее время считают, что роль Mg в поглощении света связана с тем, что он, во-первых, изменяет симметрию молекулы хлорофилла таким образом, что все сопряженные связи и сама молекула находятся в одной плоскости. Когда молекула по ка- ким-либо причинам принимает другую форму, то взаимодействие-электронных облаков нарушается, цепь сопряжения разъединяется, цвет пигментов изменяется или пропадает. Во-вторых,
83
магний придает молекуле хлорофилла способность соединяться с другими молекулами этого пигмента. И, в-третьих, магний необходим для сохранения возбужденного состояния молекулой хлорофилла. Так, тетрапиролы, содержащие вместо магния железо, быстро рассеивают свет.
Хлорофилл à — самый важный из фотосинтетических пигментов, он присутствует во всех фотосинтезирующих растениях. Существуют и другие формы хлорофиллов (b, ñ1, ñ2 è d), их называют вспомогательными (табл. 2.1; см. рис. 2.4).
|
|
|
|
|
|
|
|
|
|
|
|
|
Таблица 2.1 |
|
|
|
|
|
|
|
|
Формы хлорофилла в растениях |
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Отсутствие |
Òèï |
|
|
|
|
|
|
|
|
|
атомов Н |
|||
|
|
|
R1 |
R2 |
R3 |
R4 |
между С7 è |
||||||
хлорофилла |
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
Ñ8 (двойные |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
связи) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
à |
–ÑÍ ÑÍ2– |
–ÑÍ3 |
–ÑÍ2ÑÍ3 |
Õ |
Íåò |
||||||||
|
|
b |
–ÑÍ ÑÍ2– |
–ÑÍÎ |
–ÑÍ2ÑÍ3 |
Õ |
Íåò |
||||||
ñ1 |
–ÑÍ ÑÍ2– |
–ÑÍ3 |
–ÑÍ2ÑÍ3 |
–ÑÍ ÑÍ ÑÎÎÍ |
Äà |
||||||||
ñ2 |
–ÑÍ ÑÍ2– |
–ÑÍ3 |
–ÑÍ ÑÍ2 |
–ÑÍ ÑÍ ÑÎÎÍ |
Äà |
||||||||
|
|
d |
ÑÍÎ |
|
–ÑÍ3 |
–ÑÍ2ÑÍ3 |
Õ |
Íåò |
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Каротиноиды — жирорастворимые пигменты желтого, оранжевого и красного цветов. Они входят в состав хлоропластов и хромопластов незеленых частей растений (цветов, плодов, корнеплодов). В зеленых листьях их окраска маскируется хлорофиллом.
Каротиноиды являются тетратерпеноидами (8 остатков изопрена) и содержат 40 атомов углерода. Они представляют собой цепи, которые имеют, как и хлорофилл, двойные сопряженные связи. На одном или двух концах цепи находятся иононовые кольца.
84
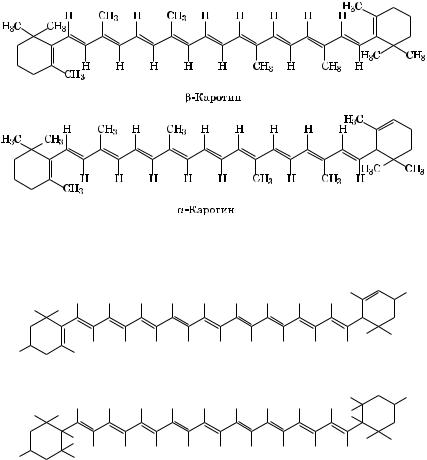
Каротиноиды делят на две группы: каротины и ксантофиллы. Каротины, например -каротин (С40Í56), представляют собой чистые углеводороды (тетратерпены):
Ксантофиллы же — лютеин С40Í56Î2 и виолаксантин С40Í56Î4 — окисленные соединения. Каротины имеют оранжевую или красную окраску, а ксантофиллы — желтую.
|
H |
CH |
H |
CH |
H |
H |
H |
H |
H C |
OH |
|
|
H |
3 |
|
||||||||
H3C |
CH3 |
3 |
|
3 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
H H H H H CH3 |
H CH3 |
H3C |
CH3 |
||||||
HO |
|
H |
|
||||||||
CH3 |
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
Лютеин |
|
|
|
|
|
|
|
H |
CH |
H |
CH |
H |
H |
H |
H |
H C |
OH |
|
|
H |
3 |
|
||||||||
H3C |
CH3 |
3 |
|
3 |
|
|
|
|
|
O |
|
|
|
|
|
|
|
|
|
|
|||
|
O |
H H H H H CH3 |
H CH3 |
H3C |
CH3 |
||||||
HO |
|
H |
|
||||||||
CH3 |
|
|
|
|
|
|
|
|
|
||
Виолаксантин
-Каротин имеет одно -иононовое кольцо (двойные связи между С5 è Ñ6), а второе — -иононовое (двойные связи между С4 è Ñ5). -Каротин отличается от -каротина тем, что имеет два-иононовых кольца.
85
Ксантофиллы отличаются большим разнообразием, чем каротины, поскольку входящие в их состав кислородосодержащие группы могут быть самыми разными: это может быть гидроксильная группа, кетогруппа, эпоксигруппа и метаксильная группа.
Распределение и типы каротиноидов в растениях имеют глубокий эволюционный смысл; их можно использовать для таксономических целей.
Относительная распространенность хлорофилла и каротиноидов в высших растениях составляет 4,5 : 1 (квантосомы содержат 230 молекул хлорофилла и 50 молекул каротиноидов). Интересно отметить, что животные обычно не синтезируют каротиноидов. Поэтому желтая и розовая расцветка птиц (например, канареек, фламинго), как и многочисленных беспозвоночных, обусловлена каротиноидами, которые они получают, поедая растения.
Молекулы каротиноидов имеют длину около 3 нм; те, которые принимают участие в фотосинтезе, обычно имеют 9 или более двойных связей.
Свет, поглощенный хлорофиллом, может вызывать образование возбужденных состояний молекулярного кислорода. Это высокореакционное состояние может повредить хлорофиллу, но взаимодействие с каротиноидами предотвращает этот процесс. Например, мутант Rhotopseudomonas spheroides, у которого отсутствуют каротиноиды, осуществляет фотосинтез в отсутствие кислорода; когда бактерии освещают в присутствии кислорода, бактериохлорофилл фотоокисляется и бактерии гибнут. У родственных штаммов, содержащих каротиноиды, такая чувствительность к кислороду не наблюдается. В других случаях, когда синтез каротиноидов подавлен, свет в присутствии кислорода может оказаться для фотосинтезирующих организмов губительным.
У дефектных по каротиноидам мутантов кукурузы и подсолнечника, а также при экспериментально нарушенном образовании каротиноидов наблюдается быстрое фотоокисление хлорофилла. Защитный (фотопротекторный) эффект объясняется способностью каротиноидов взаимодействовать с возбужденными молекулами кислорода и хлорофилла. В этом случае энергия возбужденного триплетного хлорофилла и синглетного кислорода резонансным путем передается на каротиноиды, затем рассеивается в виде тепла.
Высказывается мнение о прямом участии каротиноидов в расщеплении воды и кислородном обмене при фотосинтезе. Особое значение придается виолаксантиновому циклу:
86

При освещении в зеленых листьях наблюдается превращение виолаксантина в зеаксантин. В независимой от света реакции благодаря включению кислорода происходит обратное превращение.
В верхушках побегов растений каротиноиды обеспечивают их ориентацию к световому потоку за счет фототропизма.
Каротиноиды выполняют роль вспомогательных пигментов, которые передают энергию поглощенных квантов хлорофиллу à для совершения фотохимической работы. Особенно возрастает их значение как светоулавливающих систем в сине-фиолетовой и синей частях спектра в затененных местах, т. е. когда преобладает рассеянная радиация.
К пигментам, которые участвуют в световой стадии фотосинтеза, относятся и фикобилины. Все фотосинтетические пигменты (хлорофиллы, каротиноиды и фикобилины) входят в состав пигментных систем в виде хромпротеинов, т. е. пигмент-белковых комплексов. Хлорофиллы и каротиноиды связаны с белками относительно слабо, связь между пигментом и белком нековалентная. Поскольку такие связи легко нарушаются, хлорофиллы и каротиноиды можно экстрагировать с помощью органических растворителей (ацетон, спирт).
Фикобилины связаны с белком ковалентно, поэтому они находятся в виде молекул — фикобилипротеинов. Фикобилипротеины растворимы в воде и легко вымываются из мацерированных тканей водой или разбавленными растворами солей. Однако для разрушения молекулы фикобилипротеинов путем расщепления связи между пигментом и белком необходим гидролиз в жестких условиях.
По структуре фикобилины — тоже тетрапирроллы, но с открытой цепью, которые имеют систему конъюгированных двойных и одинарных связей. У фикобилинов четыре остатка пиррола вытянуты в открытую цепь таким образом, что замкнутого порфиринового кольца в них не образуется. В своем составе они не содержат атомов Mg или других металлов, а также фитола. Эти пигменты
87
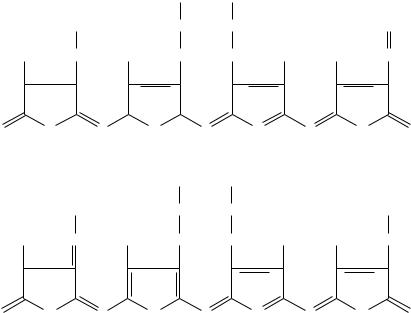
делят на две основные группы в соответствии с их цветом — красные (фикоэритрины) и синие (фикоцианины).
Название «фикобилины» предложил Лемберг (20-е гг. ХХ в.) из-за их подобия с желтыми пигментами.
Структурные формулы соответствующих билинов — фикоэритробилина (фикоэритрин), фикоцианобилина (фикоцианин), имеют следующий вид:
|
|
|
COOH |
COOH |
|
|
|
|
СH3 |
|
CH2 |
CH2 |
|
|
CH2 |
CH3 |
CH2 |
CH3 |
CH2 |
CH2 |
CH3 |
CH3 |
CH |
O |
N |
C |
N |
C |
N |
C |
N |
O |
|
H |
H |
H |
H |
|
H |
H |
|
|
|
|
|
Фикоэритрин |
|
|
|
|
|
|
|
|
COOH |
COOH |
|
|
|
|
|
С H3 |
|
CH2 |
CH2 |
|
|
CH3 |
|
CH3 |
CH |
CH3 |
CH2 |
CH2 |
CH3 |
CH3 |
CH2 |
O N C N C N C N O H H H H H H
Фикоцианин
Фикоэритрин и фикоцианин состоят из двух разных белковых субъединиц: (молекулярная масса около 19 кД) и (около 21 кД). Стехиометричные отношения - и -субъединиц составляют 1 : 1. Каждая из белковых субъединиц несет ковалентно связанный с ней фикобилин, при этом субъединицы, выделенные из разных организмов, могут нести разное количество молекул фикобилина
— от одной до четырех. Как правило, фикобилины, связанные с одной субъединицей, относятся к одному типу (т. е. к фикоэритробилинам или фикоцианобилинам); в результате получаются
88
красные или синие субъединицы. В некоторых случаях субъединица несет на себе два типа фикобилинов, но тогда один из них является преобладающим. In vivo белковая субъединица встреча- ется в виде агрегатов ( , )n, ãäå n равно 3 или 6, что соответствует молекулярным массам около 134 или 268 кД.
К фикобилинам относится и аллофикоцианин. Он назван так потому, что его сначала принимали за одну из форм фикоцианина. Аллофикоцианин присутствует в небольших количествах в водорослях семейств красных и сине-зеленых. В отличие от других фикобилипротеинов его молекула состоит только из одного типа белковых субъединиц (молекулярная масса около 15 кД), при этом каждая субъединица несет одну молекулу аллофикоцианина. In vivo она объединяется в агрегаты с молекулярной массой около 10 кД, состоящие из шести субъединиц.
Âклетках водорослей фикобилипротеины агрегируются один
ñдругим, образуя гранулы, которые называются фикобилисомами. Фикобилисомы образуют упорядоченные ансамбли на поверхности тилакоидных мембран.
Каротиноиды и фикобилины, как и хлорофиллы b, ñ1, ñ2 è d, называют вспомогательными или сопутствующими пигментами. Это определяется их функциями.
Распространение основных фотосинтетических пигментов в природе приведено в табл. 2.2.
Таблица 2.2
Распространение основных фотосинтетических пигментов в растениях
|
|
Хлорофиллы |
|
Фикобили- |
Каротиноиды |
||||||
|
|
|
протеины |
||||||||
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
Организмы |
|
|
|
|
|
|
|
Фикоэритрин |
Фикоцианин |
|
|
|
à |
|
b |
ñ1 |
ñ2 |
|
d |
Каротины |
Ксантофиллы |
||
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
Высшие |
|
|
|
|
|
|
|
|
|
|
|
растения, |
|
|
|
|
|
|
|
|
|
-Каротин |
Лютеин |
папоротнико- |
|
|
|
– |
– |
|
– |
– |
– |
Виолаксантин |
|
и мохооб- |
|
|
|
|
|
|
|
|
|
-Каротин |
Неоксантин |
разные |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Водоросли: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Лютеин |
Зеленые |
|
|
|
– |
– |
|
– |
– |
– |
-Каротин |
Виолаксантин |
|
|
|
|
|
|
|
|
|
|
|
Неоксантин |
|
|
|
|
|
|
|
|
|
|
|
|
89
|
|
|
|
|
|
|
|
|
|
Окончание табл. 2.2 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Хлорофиллы |
|
Фикобили- |
Каротиноиды |
||||||
|
|
|
протеины |
||||||||
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
Организмы |
|
|
|
|
|
|
|
Фикоэритрин |
Фикоцианин |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
à |
|
b |
ñ1 |
ñ2 |
|
d |
|
|
Каротины |
Ксантофиллы |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Неоксантин |
Эвгленовые |
|
|
|
– |
– |
|
– |
– |
– |
-Каротин |
Диодиноксантин |
|
|
|
|
|
|
|
|
|
|
|
Виолаксантин |
|
|
|
|
|
|
|
|
|
|
|
|
Бурые |
|
|
– |
|
|
|
– |
– |
– |
-Каротин |
Фукоксантин |
|
|
Виолаксантин |
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
Золотистые |
|
|
– |
|
|
|
– |
– |
– |
-Каротин |
Фукоксантин |
|
|
|
|
|
|
|
|
|
|
|
|
Желто- |
|
|
– |
– |
– |
|
– |
– |
– |
-Каротин |
Неоксантин |
зеленые |
|
|
Диодиноксантин |
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Неоксантин |
Диатомовые |
|
|
– |
|
|
|
– |
– |
– |
-Каротин |
Диодиноксантин |
|
|
|
|
|
|
|
|
|
|
|
Фукоксантин |
|
|
|
|
|
|
|
|
|
|
|
|
Криптофи- |
|
|
– |
– |
|
|
– |
|
|
-Каротин |
Аллоксантин |
товые |
|
|
-Каротин |
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
Красные |
|
|
– |
– |
– |
|
|
|
|
-Каротин |
Лютеин |
|
|
-Каротин |
Зеаксантин |
||||||||
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
Сине-зеленые |
|
|
|
|
|
|
|
|
|
|
Эхиненон |
(цианобакте- |
|
|
– |
– |
– |
|
– |
|
|
-Каротин |
Миксоксантофилл |
ðèè) |
|
|
|
|
|
|
|
|
|
|
Зеаксантин |
|
|
|
|
|
|
|
|
|
|
|
|
Если хлорофилл à встречается in vivo во всех видах растений, то b — у высших растений и двух семейств водорослей (зеленые и эвгленовые), а фикобилины только в водорослях трех семейств: красных, сине-зеленых и криптофитовых.
Спектры поглощения
Все пигменты поглощают свет избирательно. Так, если пропустить белый свет через раствор хлорофилла, а затем разложить с помощью призмы, то отдельные участки спектра окажутся сильно поглощенными и на их месте увидим черные полосы. Другие участки спектра станут проходить через раствор хлорофилла в ослабленном виде.
90
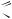
Мы получаем так называемый спектр поглощения хлорофилла. Какие длины волн будет поглощать пигмент, зависит от коли- чества и расположения двойных связей в его молекуле, от присутствия в ней ароматических колец и атомов металла. В случае хлорофилла наиболее полно поглощаются красные и сине-фиолето- вые лучи. Небольшая разница в строении молекулы хлорофиллов à è b обусловливает некоторую разницу в поглощении ими света. Хлорофилл à более полно поглощает свет с длиной волны 670, 680, 700 и 435 нм, а хлорофилл b — 650 è 480 íì.
С наименьшим поглощением проходят через раствор хлорофилла или лист зеленые лучи и часть красных. У хлорофилла b полоса поглощения в красной части спектра смещена в сторону коротковолновых лучей, а в сине-фиолетовой — в сторону длинноволновых.
Изучение спектров поглощения показало, что хлорофилл à в живом листе образует 8—10 форм, которые химически одинаковы, но отличаются по поглощению света. Как видно, такой результат обусловлен несколькими причинами. Во-первых, молекулы хлорофилла могут взаимодействовать между собой (агрегированная форма). Во-вторых, они взаимодействуют с компонентами мембран хлоропластов, в частности с белком. И, в третьих, это связано с динамическим состоянием молекул хлорофилла в тилакоидах. Хлорофилл беспрерывно разрушается под действием света (фотодеструкция). На смену разрушенным молекулам в мембраны тилакоидов встраиваются новые. Некоторые особенности окраски пигментов и спектры поглощения представлены в табл. 2.3.
|
|
|
|
|
Таблица 2.3 |
|
|
Некоторые особенности пигментов |
|
||||
|
|
|
|
|
|
|
|
Структурные |
Отличия в спектрах |
|
|||
Пигменты |
поглощения; |
Окраска |
||||
|
|
отличия |
||||
|
|
|
длина волны, нм |
|
||
|
|
|
|
|
||
|
|
|
|
|
|
|
Хлорофилл à |
–ÑÍ3 |
670, 680, 700, 435 |
Сине-зеленая |
|||
|
|
|
|
|
|
|
|
|
|
O |
|
|
|
Хлорофилл b |
|
|
C |
650, 480 |
Желто-зеленая |
|
|
|
|||||
|
|
|
H |
|
|
|
|
|
|
|
|
|
|
-Каротин |
Ñ40Í56 |
425, 450, 480 |
Желтая |
|||
Лютеин |
Ñ40Í56Î2 |
425, 445, 450 |
Желтая |
|||
91