

Fiziologia_rasteny
.pdfФотосистема — это совокупность светособирающего комплекса (ССК), фотохимического реакционного центра и связанных с ним молекул — переносчиков электрона. По современным представлениям фотосистема является важнейшим структурно-функ- циональным звеном фотосинтетического аппарата. Две пигментные фотосистемы — это две машины, движущие фотосинтез на световой стадии.
Каждой фотосистеме соответствует отдельный набор пигментов, а также связанных с ними переносчиков электронов, и в каждой фотосистеме происходят свои, только ей присущие, фотосинтетические реакции.
Из анализа работ следует, что в тилакоидных мембранах хлоропластов формируются пигмент-белковые комплексы, вклю- чающие фоторецепторные системы, что имеет решающее значе- ние для осуществления основных функций пигментов в фотосинтезе.
Одной из функций является организация фотосинтетического аппарата в виде светособирающих комплексов, функционально сопряженных с реакционным центром для обеспечения коллективного и эффективного поглощения фотонов и передаче ее в реакционные центры.
Âреакционных центрах энергия электронного возбуждения используется для фотохимических реакций.
Âсостав светособирающих пигментов (светособирающие антен-
ны) входят: каротин, Хл à662, à670, à677, à684, à692 (ÔÑ I) è ÔÑ II: à622, à670, à677. Кроме того, имеется еще светособирающий белко-
вый комплекс (ССК) (рис. 2.8).
Считают, что при низкой интенсивности света собственные антенны ФС I и ФС II поглощают мало квантов света и поэтому не могут поддерживать необходимой скорости фотосинтезирующих реакций, нужен дополнительный приток энергии, который обеспечит ССК. Наоборот, при высоких интенсивностях света пигментным системам не нужно столько энергии и поэтому они содержат значительно меньше ССК.
В состав ССК входят ксантофиллы, хлорофилл b, некоторое количество хлорофилла à (см. рис. 2.8). ССК передает поглощенную энергию на реакционный центр (рис. 2.9).
Получив эту энергию, специальная форма хлорофилла в реакционном центре переходит в возбужденное состояние (Хл*). Õë* обладает очень высокой реакционной способностью, это сильный восстановитель.
112

ССП
ФС I Каротины  Хл а662
Хл а662  а670
а670  а677
а677  а684
а684  а692
а692 
 Р700
Р700
ФС II |
Хл а662 |
|
а670 |
|
а677 |
|
|
|
Р680 |
|
||||
|
|
|
|
|
|
|||||||||
|
|
|
|
|
|
ССК |
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
Ксантофиллы |
|
|
Хл b (Хл c) |
|
|
|
Хл a |
||||||
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Ðèñ. 2.8. Схема организации ФС I и ФС II
ССК Ксантофиллы |
Хл b (Хл c) |
Хл a |
ССП |
|
ССП |
ФС II |
ФС I |
|
P680 |
|
P700 |
H2O |
|
НАДФ+ + Н+ |
e |
|
e |
1/2 O2 |
|
НАДФ |
Ðèñ. 2.9. Схема взаимодействия пигментов ФС I, ФС II
èсветособирающего комплекса
Âокислительно-восстановительной системе А/А– происходит восстановление А до А–. Îò À– электрон переходит дальше и через
другие компоненты электрон-транспортной цепи переносятся на НАДФ .
Хлорофилл, который отдал электрон, переходит в состояние свободного катиона — радикала Х . Он возвращается в основное первичное состояние, получив электрон восстановленной фор-
113

ìû Ä– — второй окислительно-восстановительной системы Д/Д–. Окисленный Д восстанавливается до Д–, получая электрон от воды (рис. 2.10).
Ðèñ. 2.10. Упрощенная схема работы пигментной системы
Догадку о том, что кислород должен выделяться из воды, высказал в 1931 г. К. Ван-Нил (Голландия). В 1937 г. англичанин Р. Хилл показал, что изолированные хлоропласты выделяют на свету кислород в условиях наличия акцептора электронов (феррицианид, бензохинон и др.). Реакцию Р. Хилла в общем в виде записывают так:
Реакция разложения воды происходит в три этапа. В данном случае акцептором является хинон. Таким образом, процессы выделения О2 и восстановления СО2 независимы один от другого.
Â1941 г. одновременно две группы ученых: А. П. Виноградов
èР. В. Тейс в СССР и С. Рубен и М. Д. Камен в США, доказали, что весь кислород, который выделяют растения при фотосинтезе,
является кислородом воды, а не СО2, как считали раньше.
ФС II и ФС I размещаются последовательно в цепи транспорта
электронов от Н2О до НАДФ . Для восстановления одной молекулы НАДФ в процессе фотосинтеза необходимо два электрона и два протона, при этом донором электронов является вода. Фото-
114

индуцированное окисление воды происходит в ФС II, восстановление НАДФ — ФС I. Таким образом, фотосистемы должны функционировать во взаимодействии.
Исходя из данных квантового выхода фотосинтеза (количество выделенного О2 или связанного СО2 на 1 квант поглощенной энергии), необходимо восемь квантов света для выделения одной молекулы О2; из состава компонентов ФС I и ФС II и величины окислительно-восстановительного потенциала была разработана схема последовательности реакций в световой стадии фотосинтеза. Из-за подобия с буквой Z эта схема получила название Z-схемы (рис. 2.11).
Впервые принцип Z-схемы был предложен Р. Хиллом и Ф. Бендаллом (1960) и экспериментально подтвержден работами Л. Дюйзенса (1961).
Ðèñ. 2.11. Z-схема переноса электронов в фотохимических реакциях у фотосинтетических организмов
ФС I — единственная, имеющаяся у бактерий, работает без участия кислорода; эта ФС, вероятно, имела преимущества на ранних этапах биологической эволюции, когда кислорода в земной атмосфере было мало. С развитием ФС II растения получили возможность выделять молекулярный кислород из воды. Вероятно, это обстоятельство и определило изменения свойств земной атмосферы: из анаэробной она стала аэробной.
115
Таким образом, фотохимическую работу у высших растений, выполняемую при фотосинтезе, в конечном счете можно свести до разложения воды.
Однако сначала физическая энергия уловленных фотонов должна быть видоизменена и переведена в «химическую» энергию. Мы отметили, что энергия мигрирует до реакционных центров. В результате отдельные электроны получают достаточное количество энергии для того, чтобы они могли перейти от молекул хлорофилла Р700 èëè Ð680 до молекул — переносчиков электронов. Переносчик поглощает определенную часть этой энергии активации и передает электрон следующему переносчику, где этот процесс повторяется. В хлоропластах разные переносчики электронов расположены на мембране или внутри мембран и образуют ряд, в пределах которого они располагаются в соответствии со способностью присоединять электроны (другими словами,
ñèõ окислительно-восстановительным потенциалом). Таким образом, электрон переходит от одного переносчика к другому, подобно тому, как движется по каскаду вода — с одного уступа на другой.
Поясним суть основных элементов, входящих в ФС I и ФС II. Реакционный центр ФС II включает кислородовыделяющий
комплекс, хлорофилл Р680, первичный акцептор электронов феофитин (Фео) — производное Хл, в котором Mg замещен протона-
ми, и вторичные акцепторы — молекулы монохинона (QÀ è QÂ), а также пластохинон (Q).
Ядро реакционного центра ФС II составляют два мембранных белка с ММ 32 и 34 кД. Оба белка имеют по пять трансмембранных петель и служат основой большинства простетических групп, выполняющих функции переносчиков электронов. Другие белки (43 и 47 кД) ФС II входят в состав комплекса светособирающих пигментов (ССП) или участвуют в выделении кислорода при фотоокислении воды (33, 23, 16 кД). Функции остальных белков
ñнебольшой молекулярной массой, например цитохрома b559 и других, пока неизвестны.
Âкислородовыделяющий комплекс входят Mn-содержащий пул (кластер) и как кофакторы кальций и хлор. Посредником ме-
жду кислородовыделяющим центром и хлорофиллом Р680 является остаток аминокислоты тирозина (Yz).
Реакционный центр ФС I включает первичный донор электронов Хл Р700, первичный акцептор Хл à (À0), акцепторы — же-
116
лезосерные белки (Fe4S4, Fõ, FÀ, FÂ), водорастворимый железосерный белок (Fe2S2), белок ферредоксин (ФÄ), растворимый флавопротеин — ферредоксин-НАДФ-редуктазу (Фï).
Комплекс ФС I представляет собой интегральный пигмент-бел- ковый комплекс с ММ около 340 кД. Он включает 13 пептидов, из которых 8 — интегральных и 5 — периферических. К комплексу примыкают белки внешней антенны ССК I. Интегральные белки имеют массу 83,0 и 82,4 кД соответственно, а белок с ММ 8,9 кД расположен на поверхности мембраны со стороны стромы. Первые два белка присоединяют димер Р700, по одной молекуле мономера Хл à695, по одной молекуле филлохинона, а также железосерный кластер Fx. Белок с ММ 8,9 кД вместе с тремя низкомолекулярными белками образуют периферийный домен со стороны стромы. Он содержит железосерные кластеры с центрами FÀ è FÂ. Белки 9,0 и 15,7кД составляют периферийный домен и связывают восстановленный пластоцианин с ФС I.
Комплекс «цитохром b6 /f» состоит из цитохрома b6, цитохрома f, железосерного белка Риске (Fe2S2). Цитохром b6 является гидрофобным элементом комплекса.
Простетическая группа цитохрома f представлена гемом ñ-типа, а аналогичная группа цитохрома b6 состоит из двух гемов b-типа, ковалентно связанных с остатками гистидина. Один гем называют высокопотенциальным (Н), второй — низкопотенциальным (L).
В передаче электронов от ФС II, а именно от комплекса b6 /f к ФС I участвует пластоцианин (водорастворимый белок, содержащий два атома меди).
Сущность реакций, происходящих на свету, была выяснена Д. Арноном (США) в 1954—1958 гг. Он показал, что на свету в изолированных хлоропластах идет образование АТФ за счет присоединения минерального фосфора (Ф) к АДФ.
Во время перемещения электрона по такой цепи переносчи- ков часть его энергии преобразуется в химическую форму, поскольку за счет нее из АДФ и неорганического фосфора синтезируется АТФ и запасается значительное количество энергии — 8—10 ккал/моль.
Дальнейшие исследования доказали, что синтез АТФ сопряжен с транспортом электронов — циклическим или нецикличе- ским.
117

Циклический транспорт электронов
В первой пигментной системе пигментом-ловушкой является Р700, а остальные — сборщики (антенный комплекс). При поглощении кванта света один из электронов Р700 переходит на более высокий энергетический уровень и присоединяется к Хл à695, а затем передается на белок, содержащий железо и серу (FeS). Окислительно-восстановительный потенциал (Е) Р700 в основном состоянии составляет 0,43 В; таким образом, тенденция к потере электронов у него выявлена слабо. Когда в результате поглощения кванта света молекула Р700 переходит в возбужденное состояние, ее окислительно-восстановительный потенциал падает до –0,6 В, и поэтому она легко отдает электрон (рис. 2.12).
Ðèñ. 2.12. Упрощенная схема циклического фотофосфорилирования
От белка, содержащего железо и серу, электрон передается переносчику ферредоксину (ФД). От ферредоксина электрон вновь возвращается к Р700 через ряд промежуточных переносчиков, среди которых цитохромы b6 è f, а также пластоцианин (Пц). В то время, когда электрон транспортируется по этому циклическому пути, его энергия используется для присоединения Фí к АДФ с образованием АТФ.
Нециклический транспорт электронов
Когда устьица листа открыты, а хлоропласты поглощают свет разных длин волн, одновременно с ФС I включается ФС II. Совместно они составляют систему нециклического транспорта электронов.
118

Как ранее отмечалось, под действием света Р700 переходит в возбужденное состояние (Р700*) и окисляется (Р700 ), электроны переносятся на акцепторы А0 (хлорофилл à) è À1 (филлохинон). Далее на мембранно-связанные железосерные белки (Fx, FÀ, FÂ), через которые электроны поступают на ферредоксин. Но в этом слу- чае от последнего через флавопротеин — ферредоксин-НАДФ-ре- дуктазу — электроны идут на восстановление НАДФ до НАДФН (см. рис. 2.11).
Когда два электрона присоединяются к НАДФ из водной среды, которая окружает тилакоидные мембраны, к электрону присоединяются протоны воды и образуется НАДФН, который используется в последующих реакциях для восстановления СО2 до уровня углевода.
Таким образом, Р700 окислен, в нем образовалась электронная «дырка». Он не может долго оставаться в этом состоянии. Откуда он получает электрон?
При поглощении кванта света Р680 тоже переходит в возбужденное состояние и отдает электрон. При этом Хл Р680 переходит в окисленную форму (Р 680), а феофитин восстанавливается (Фео–). Затем электрон передается на два хинона QA è QB (монохиноны). После этого электроны поступают на липидрастворимый пул пластохинонов (Q), который функционирует как «двухэлектронные ворота». Один из электронов поступает в цикл цитохромов b6 (Öèò b/f), а второй направляется по линейной цепи в ФС I. Затем электроны переносятся на водорастворимый белок пластационин (Пц) и в ловушку Р700. В результате восстанавливается окисленная форма хлорофилла Р 700.
Но теперь «дырка» появилась на Р680. Она заполняется присоединением электрона от воды. Электрон под воздействием света в результате фотоокисления (фотолиз) поступает от молекулы воды. В результате молекула воды разрушается до протонов и кислорода:
2H2O свет 4e + 4H+ + O2 .
4e + 4H+ + O2 .
Ингибитором этой реакции является диурон (дихлорфенилдиметилмочевина). Растения, обработанные диуроном, гибнут, так как при этом блокируется транспорт электронов от воды, не образуется АТФ и не восстанавливается НАДФ , а следовательно, невозможно восстановление СО2.
Образованные при фотоокислении воды протоны остаются в водной среде тилакоидов хлоропластов, которая включает не только
119
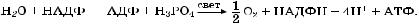
молекулы воды, но также Н - и ОН–-ионы. Накопление протонов на внутренней стороне мембраны тилакоидов играет определенную роль в образовании АТФ, о чем разговор пойдет дальше. Этот путь транспорта электронов получил название нециклического.
Нециклический транспорт электронов также связан с синтезом АТФ. Процесс фосфорилирования АДФ с образованием АТФ, связанный с нециклическим транспортом электронов, получил название нециклического фотосинтетического фосфорилирования (см. рис. 2.11).
Таким образом, процессы световой фазы фотосинтеза в общем виде можно записать
Во время световой фазы происходит перенос электронов от воды (Е 0,81 В) на НАДФ (Е –0,32 В). Перенос электронов от вещества с более положительным потенциалом к веществу с более отрицательным требует затраты энергии. Используется энергия поглощенного пигментом света. Для транспорта каждого электрона требуется два кванта. Этот перенос электронов против градиента окислительно-восстановительного потенциала и является фотохимической работой.
Следовательно, сущность световой реакции фотосинтеза — вызванный светом последовательный транспорт электронов от одной молекулы к другой, в результате образуются АТФ и НАДФН (рис. 2.13).
Так как молекула, которая электрон теряет, окисляется, а которая получает — восстанавливается, то это окислительно-вос- становительный процесс. Для транспорта электронов от одного переносчика ко второму, как и для передачи энергии между молекулами пигментов, необходима тщательная упаковка молекул и строгая упорядоченность в их размещении, поэтому световая стадия фотосинтеза связана с тилакоидными мембранами хлоропластов. Кроме этого, с помощью мембран происходит разобщение процессов образования кислорода и мощного восстановителя, в противном случае, они могли бы взаимодействовать. Синтез АТФ идет в строме.
Основные различия этих двух видов фосфорилирования даны в табл. 2.4.
Следовательно, в циклическом транспорте электронов участвует только ФС I, в нециклическом — две. ФС I расположена в тилакоидах стромы, а ФС II — только в тилакоидах гран.
120

Ðèñ. 2.13. Локализация электрон-транспортных реакций в мембране тилакоида
|
|
Таблица 2.4 |
|
Сравнение циклического и нециклического |
|||
фотофосфорилирования |
|
||
|
|
|
|
Процесс |
Нециклическое |
Циклическое |
|
|
|
|
|
Путь электронов |
Нециклический |
Циклический |
|
|
|
|
|
Первый донор (источник) |
Âîäà |
ÔÑ I (Ð700) |
|
электронов |
|||
|
|
||
|
|
|
|
Последний акцептор (место |
ÍÀÄÔ |
ÔÑ I (Ð700) |
|
назначения) электронов |
|
|
|
|
|
|
|
|
Основные: |
Основные: |
|
Продукты |
АТФ, НАДФН |
ÀÒÔ |
|
|
Побочные: О2 |
|
|
Участвующие ФС |
I è II |
I |
|
|
|
|
|
Электроны, поступающие от ферредоксина, могут также переноситься и восстанавливать кислород с образованием Н2Î2 и, в конечном счете, Н2О. В этом случае функционируют ФС I и ФС II и поглощаются с использованием восстановленного ферредоксина в особой реакции равные количества кислорода; такой поток электронов получил название псевдоциклического. Хотя при этом потоке электронов отсутствует суммарное изменение со-
121