

Ярыгин_Биология_кн_1_2004
.pdfРис. 3.49. Структурные блоки в организации хрома тина. А —петельная структура хроматина; Б —даль нейшая конденсация хроматиновых петель; В — объединение петель, имеющих сходную структуру, в блоки с образованием окончательной формы интер фазной хромосомы
Примером факульта тивного гетерохроматина служит тельце полового хроматина, образуемое в норме в клетках организ мов гомогаметного пола (у человека гомогаметним является женский пол) одной из двух Х-хромо- сом. Гены этой хромосо мы не транскрибируются. О бразование факульта тивного гетерохроматина за счет генетического ма териала других хромосом сопровождает процесс клеточной дйфференцировки и служит механиз мом выключения из активной функции групп генов, транскрипция ко торых не требуется в клет ках данной специализа ции. В связи с этим рису нок хроматина ядер клеток из разных тканей и органов на гистологиче ских препаратах различа ется. Примером может служить гетерохроматизация хроматина в ядрах зрелых эритроцитов птиц.
|
Перечисленные уров |
|||||
|
ни структурной организа |
|||||
|
ции |
|
хроматина |
обна |
||
|
руживаются в неделящей- |
|||||
|
ся клетке, когда хромосо |
|||||
|
мы |
еще |
недостаточно |
|||
|
компактизованы, чтобы |
|||||
|
быть видимыми в свето |
|||||
Ю МКМ •"Ji |
вой |
микроскоп |
как от |
|||
дельные структуры. Лишь |
||||||
некоторые |
их участки с |
|||||
|
более |
высокой |
плотно |
|||
|
стью упаковки выявляют |
|||||
|
ся |
в |
ядрах |
в |
виде |
|
Рис. 3.50. Конститутивный гетерохроматин в метафаз- |
хроматиновых |
|
глыбок |
|||
ных хромосомах человека |
(рис. 3.51). |
|
|
|
||
Рис. 3.51. Гетерохроматин в интерфазном ядре Компактные участки гетерохроматина сгруппированы около ядрышка и ядерной мембраны
Метафазная хромосома. Вступление клетки из интерфазы в митоз сопровождается суперкомпактизацией хроматина. Отдельные хромо сомы становятся хорошо различимы. Этот процесс начинается в профазе, достигая своего максимального выражения в метафазе митоза и анафазе (см. разд. 2.4.2). В телофазе митоза происходит декомпактизация вещества хромосом, которое приобретает структуру интерфаз ного хроматина. Описанная митотическая суперкомпактизация облегчает распределение хромосом к полюсам митотического веретена
ванафазе митоза. Степень компактизации хроматина в разные периоды митотического цикла клетки можно оценить по данным, приведенным
втабл. 3.2.
3.5.2.3. Морфология хромосом
Митотическая суперкомпактизация хроматина делает возможным изучение внешнего вида хромосом с помощью световой микроскопии. В первой половине митоза они состоят из двух хроматид, соединенных между собой в области первичной перетяжки (центромеры или тнетохора) особым образом организованного участка хромосомы, общего для обеих сестринских хроматид. Во второй половине митоза проис ходит отделение хроматид друг от друга. Из них образуются однонитчатые дочерние хромосомы, распределяющиеся между дочерними клетками.
В зависимости от места положения центромеры и длины плеч, расположенных по обе стороны от нее, различают несколько форм хромосом: равноплечие, или метацентрические (с центромерой посе редине), неравноплечие, или субметацентрические (с центромерой, сдвинутой к одному из концов), палочковидные, или акроцентриче-
122 |
123 |
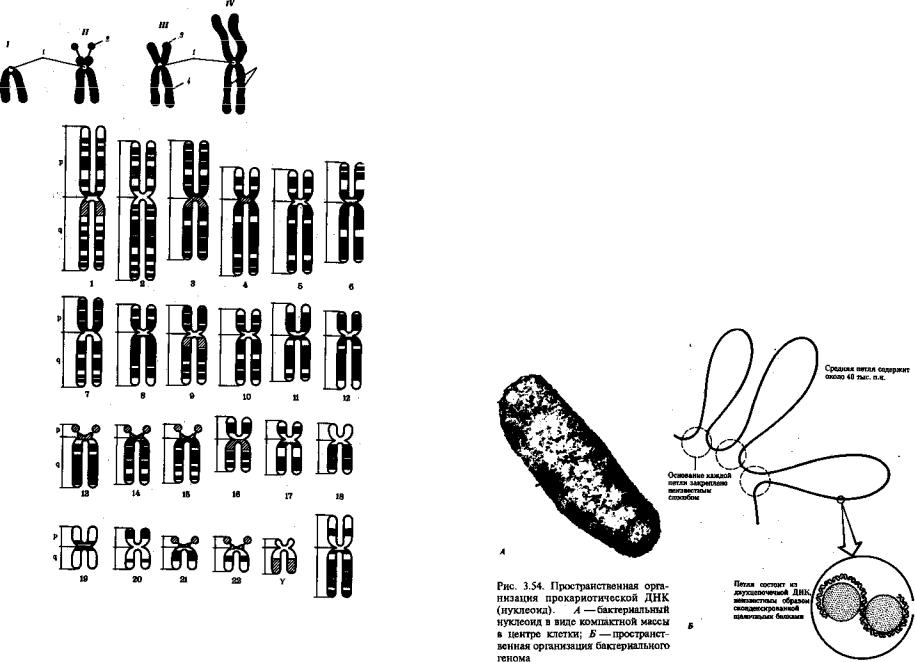
Рис. 3.52. Формы хромосом: /—телоцентрическая, //—акро- центрическая, ///—субметацент- рическая, IV—метацентрическая; 1 —центромера, 2 —спутник, 3 — короткое плечо, 4— длинное пле-
5 чо,5—хроматиды
Рис. 3.53. Расположение локусов в хромосомах человека при их дифференциальном окрашивании:
р — короткое плечо, q — длинное плечо; 1—22 — порядковый номер хромосомы; XY — половые , |
|
хромосомы |
і |
ские (с центромерой, расположенной практически на конце хромосо мы), и точковые —очень небольшие, форму которых трудно опреде лить (рис. 3.52). При рутинных методах окраски хромосом они различаются по форме и соотносительным размерам. При использо вании методик дифференциальной окраски выявляется неодинаковая флуоресценция или распределение красителя по длине хромосомы, строго специфические для каждой отдельной хромосомы и ее гомолога (рис. 3.53).
Таким образом, каждая хромосома индивидуальна не только по заключенному в ней набору генов, но и по морфологии и характеру дифференциального окрашивания.
3.5.2.4. Особенности пространственной организации генетического материала в прокариотической клетке
Описанная организация наследственного материала эукариот, свя занная с расположением его функциональных единиц в ядерных структурах — хромосомах, у прокариот отсутствует. Основная часть генетического материала заключена у них в единственной кольцевой молекуле ДНК длиной около 1 мм (Е. coli), причем у прокариот не обнаружено гистонов, обеспечивающих нуклеосомную организацию хроматина эукариот. Однако кольцевой молекуле ДНК прокариот
125

также присуща укладка в виде петель. ДНК прокариот тоже комплексируется с некоторыми негистоновыми белками, образуя нуклеоид, который гораздо менее устойчив по сравнению с хроматином эукариот (рис. 3.54).
3.5.3. Проявление основных свойств материала наследственности и изменчивости
на хромосомном уровне его организации
На хромосомном уровне организации, который появляется в про цессе эволюции у эукариотических клеток, генетический аппарат должен удовлетворять всем требованиям, предъявляемым к субстрату наследственности и изменчивости: обладать способностью к самовос произведению, поддержанию постоянства своей организации и при обретению изменений, которые могут передаваться новому поколению клеток.
3.5.3.1. Самовоспроизведение хромосом в митотическом цикле клеток
Процесс самовоспроизведения —редупликации хромосом осуще ствляется в период подготовки клетки к митозу —в интерфазе. Вслед ствие этого в ходе последующего деления клетка передает дочернему
|
поколению полноценный ком |
||||
|
плекс |
генов, |
заключенный в |
||
|
каждой отдельной хромосоме. |
||||
|
В основе |
самовоспроизведе |
|||
|
ния хромосомы лежит репли |
||||
|
кация ДНК с образованием из |
||||
|
одной |
материнской спирали |
|||
|
двух дочерних молекул, на базе |
||||
|
которых образуются две |
сест |
|||
|
ринские хроматиды (рис. 3.55). |
||||
|
Благодаря |
высокой точности |
|||
|
репликации ДНК (см. разд. |
||||
|
3.4.2.2 ) сестринские хромати |
||||
|
ды несут практически одина |
||||
|
ковую информацию, записан |
||||
|
ную в ее дочерних молекулах. |
||||
|
В каждой хроматиде ДНК на |
||||
|
ходится в комплексе с гисто- |
||||
Рис. 3.55. Возникновение двунипатой хромо- |
н а м и > |
и |
е Й |
присущи |
все |
сомы из однонитчатой: |
ОПИСаННЫе ВЫШе урОВИИ КОМ- |
||||
/—интерфазная хромосома до репликации ДНК, ПаКТИЗаЦИИ ХрОМаТИНа, СВОЙ- II — интерфазная хромосома после репликации ДНК, СТВЄННЬІЄ ИНТЄрфаЗНОЙ КЛЄТКЄ.
///— метафазная хромосома
Нормальные митотические хромосомы при том
же масштабе увеличения
Левое плечо хромосомы 3
Рис. 3.56. Полный набор политенных хромосом из клетки слюнной железы дрозофилы
Таким образом, готовясь к самовоспроизведению, клетка удваивает содержание ДНК в каждой хромосоме. При этом последняя приобре тает двунитчатую структуру.
Известны случаи, когда репликация ДНК, причем неоднократная, не сопровождается формированием хроматид на базе дочерних спира лей. При этом образуются так называемые полишенные хромосомы, содержащие многие сотни копий ДНК. Такие хромосомы обнаружи ваются, например, в неделящихся клетках слюнных желез личинок неко торых насекомых при обычной световой микроскопии (рис. 3.56).
126 |
127 |

3.5.3.2. Распределение материала материнских хромосом между дочерними клетками в митозе
В ходе митотического деления обеспечивается закономерное рас пределение сестринских хроматид каждой хромосомы между дочерни ми клетками. В составе дочерних хромосом (бывших сестринских хроматид) каждая клетка нового поколения получает одну из двух молекул ДНК, образовавшихся в результате репликации материнской двойной спирали. Следовательно, новое поколение клеток получает одинаковую генетическую информацию в составе каждой группы сцепления.
Таким образом, процессы, происходящие с хромосомами при подготовке клеток к делению и в самом делении, обеспечивают самовоспроизведение и постоянство их структуры в ряду клеточных поколений (см. разд. 3.6.2.1).
После митоза хромосомы дочерней клетки представлены одной молекулой ДНК, компактно упакованной с помощью белков в одну хроматиновую нить, т.е. имеют такую же структуру, какую имели хромосомы материнской клетки до начала процесса репликации ДНК. Если вновь образованная клетка выбирает путь подготовки к делению, то в ней должны произойти все описанные выше события, связанные с динамикой структурной организации ее хромосом.
3.5.3.3. Изменения структурной организации хромосом. Хромосомные мутации
Несмотря на эволюционно отработанный механизм, позволяющий сохранять постоянной физико-химическую и морфологическую орга низацию хромосом в ряду клеточных поколений, под влиянием раз личных воздействий эта организация может изменяться. В основе изменения структуры хромосомы, как правило, лежит первоначальное нарушение ее целостности — разрывы, которые сопровождаются раз личными перестройками, называемыми хромосомными мутациями или
аберрациями.
Разрывы хромосом происходят закономерно в ходе кроссинговера, когда они сопровождаются обменом соответствующими участками между гомологами (см. разд. 3.6.2.3). Нарушение кроссинговера, при котором хромосомы обмениваются неравноценным генетическим ма териалом, приводит к появлению новых групп сцепления, где отдель ные участки выпадают — делеции—или удваиваются—дупликации (рис. 3.57). При таких перестройках изменяется число генов в группе сцепления.
Разрывы хромосом могут возникать также под влиянием различных мутагенных факторов, главным образом физических (ионизирующего и других видов излучения), некоторых химических соединений, вирусов.
128
А В С |
D |
E F G H |
|
А В Г Р Р Г Н |
А В С |
D |
Е F G H |
_ |
A B C B C D E F G H |
А В С D Е |
F G Н Перицентрическая инверсия A B C |
G |
Н |
||||
|
|
|
|
|
|
||
VI і ЩУ&ІР- |
-СЕЕШСШЮ |
||||||
f |
|
f |
|
|
F E D |
|
|
A B O D E |
|
F G HПарацентрическая инверсия A |
E F |
G H |
|||
СЖШХГО - С Ж Ж Х Ш
t |
t |
D С В |
||
A B C D E F G H |
M N O C D E F G H |
|||
( I I I I-XTD |
^ Ш Х Ш |
|||
f |
Реципрокная транслокация |
|||
|
• |
— |
|
» . |
M N O P Q R |
A B P Q R |
|||
|
t |
|
||
А В С |
D E F G н Н е Р е ц и п Р ° к н а я транслокацияА D E F В С G H |
|||
• . • г
Рис. 3.57. Виды хромосомных перестроек
Нарушение целостности хромосомы может сопровождаться пово ротом ее участка, находящегося между двумя разрывами, на 180° — инверсия. В зависимости от того, включает ли данный участок область центромеры или нет, различают перицентпрические и парацентрические инверсии (рис. 3.57).
Фрагмент хромосомы, отделившийся от нее при разрыве, может быть утрачен клеткой при очередном митозе, если он не имеет центромеры. Чаще такой фрагмент прикрепляется к одной из хро мосом — транслокация. Нередко две поврежденные негомологичные хромосомы взаимно обмениваются оторвавшимися участками —ре ципрокная транслокация (рис. 3.57). Возможно присоединение фраг мента к своей же хромосоме, но в новом месте — транспозиция (рис. 3.57). Таким образом, различные виды инверсий и транслокаций характеризуются изменением локализации генов.
Хромосомные перестройки, как правило, проявляются в изменении морфологии хромосом, что можно наблюдать в световой микроскоп. Метацентрические хромосомы превращаются в субметацентрические и акроцентрические и наоборот (рис. 3.58), появляются кольцевые и полицентрические хромосомы (рис. 3.59). Особую категорию хромо-
9. Ярыгин. Т. 1. |
129 |

|
|
|
|
/^~^\ |
|
|
|
|
Рис. 3.58. Изменение формы хро- |
|
|
А . В . С |
e e j * |
G |
" |
|
м |
мосом в результате перицентри- |
|
|
X |
І |
І |
ттЖЯ |
I |
|
> |
Мантрическая |
ческИХ ИНВерСИЙ |
|
|
A R C " |
F E |
П |
С |
H |
|
|
|
\ |
і |
і |
тжтж і |
|
> |
СУ*-—п—« |
|
||
|
|
|
|
|
|
|
|
|
|
A |
B |
C |
D |
|
E |
F G |
H |
|
|
с |
і |
|
і |
в а |
Ч |
Метацентрическая |
|||
A |
B |
C |
G |
F E |
D |
Н |
|
||
Сгтт |
|
|
|
|
yjfotfffiX """^ |
Акроцентрическая |
|||
|
|
|
|
А |
В |
|
C D |
E F |
G H |
'•ХГХП
|
А |
В |
C D |
E F G H |
|
« ( |
|
I I |
I |
К П П |
|
|
К |
L |
М N |
|
О Р |
A B C D E F G L M N O P
г т - т - г т і і і і —і—і—«—і—>
Рис. 3.59. Образование кольцевых (I) и полицентрических (II) хромосом
А В С D Е F G
Н И И IXD
МN О Р Q R
Рис. 3.60. Хромосомные перестройки, связанные с центрическим слиянием или разде лением хромосом Являются причиной изменения числа хромосом в кариотипе
сомных мутации представляют аберрации, связанные с цент рическим слиянием или разде лением хромосом, когда две негомологичные структуры объединяются в одну — робертсоновская транслокация,
или одна хромосома образует две самостоятельные хромосо мы (рис. 3.60). При таких мута циях не только появляются хромосомы с новой морфоло гией, но и изменяется их коли чество в кариотипе.
Описайные структурные изменения хромосом, как пра вило, сопровождаются измене нием генетической программы,
Рис. 3.61. Петля, образующаяся при конъюга ции гомологичных хромосом, которые несут неравноценный наследственный материал в соответствующих участках в результате хромо сомной перестройки
получаемой клетками нового поколения после деления материнской клетки, так как изменяется количественное соотношение генов (при делециях и дупликациях), меняется характер их функционирования в связи с изменением взаимного расположения в хромосоме (при ин версии и транспозиции) или с переходом в другую группу сцепления (при транслокации). Чаще всего такие структурные изменения хромо сом отрицательно сказываются на жизнеспособности отдельных сома тических клеток организма, но особенно серьезные последствия имеют хромосомные перестройки, происходящие в предшественниках гамет.
ІЙ
ABCJ
я — >
ABCji (Р
Сі
Рис. 3.62. Образование
при конъюгации квадривалента из двух пар хромосом, несущих реципрокную трансло кацию
д*
ш
<ж
Рис. 3.63. Образование при конъюгации поливалента шестью парами хромосом, участвующих в реципрокных транслокациях:
I—конъюгация между парой хромосом, не несущих транслокацию; /7—поливалент, образуемый шестью парами хромосом, участвую щих в транслокации
131

Изменения структуры хромосом в предшественниках гамет сопро вождаются нарушением процесса конъюгации гомологов в мейозе и их последующего расхождения. Так, делеции или дупликации участка одной из хромосом сопровождаются при конъюгации образованием петли гомологом, имеющим избыточный материал (рис. 3.61). Реципрокная транслокация между двумя негомологичными хромосомами приводит к образованию при конъюгации не бивалента, а квадривалента, в котором хромосомы образуют фигуру креста благодаря при тягиванию гомологичных участков, расположенных в разных хромосомах (рис. 3.62). Участие в реципрокных транслокациях боль шего числа хромосом с образованием поливалента сопровождается формированием еще более сложных структур при конъюгации (рис. 3.63).
В случае инверсии бивалент, возникающий в профазе I мейоза, образует петлю, включающую взаимно инвертированный участок (рис.
Конъюгация и последующее расхождение структур, образованных измененными хромосомами, приводит к появлению новых хромосом ных перестроек. В результате гаметы, получая неполноценный наслед ственный материал, не способны обеспечить формирование нормального организма нового поколения. Причиной этой является нарушение соотно шения генов, входящих в состав отдельных хромосом, и их взаимного расположения.
Однако, несмотря на неблагоприятные, как правило, последствия хромосомных мутаций, иногда они оказываются совместимыми с жизнью клетки и организма и обеспечивают возможность эволюции структуры хромосом, лежащей в основе биологической эволюции. Так, небольшие по размеру делеции могут сохраняться в гетерозиготном состоянии в ряду поколений. Менее вредными, чем делеции, являются
Рис. 3.64. Конъюгация хромосом при инверсиях:
/ — парацентрическая инверсия в одном из гомологов, II—перицентрическая инверсия в одном из гомологов
дупликации, хотя большой объем материала в увеличенной дозе (более 10% генома) приводит к гибели организма.
Нередко жизнеспособными оказываются робертсоновские транс локации, часто не связанные с изменением объема наследственного материала. Этим можно объяснить варьирование числа хромосом в клетках организмов близкородственных видов. Например, у разных видов дрозофилы количество хромосом в гаплоидном наборе колеб лется от 3 до 6, что объясняется процессами слияния и разделения хромосом. Возможно, существенным моментом в появлении вида Homo sapiens были структурные изменения хромосом у его обезьяноподоб ного предка. Установлено, что два плеча крупной второй хромосомы человека соответствуют двум разным хромосомам современных чело векообразных обезьян (12-й и 13-й—шимпанзе, 13-й и-14-й—го риллы и орангутана). Вероятно, эта человеческая хромосома образовалась в результате центрического слияния по типу робертсоновской транслокации двух обезьяньих хромосом.
Ксущественному варьированию морфологии хромосом, лежащему
воснове их эволюции, приводят транслокации, транспозиции и ин версии. Анализ хромосом человека показал, что его 4, 5, 12 и 17-я хромосомы отличаются от соответствующих хромосом шимпанзе перицентрическими инверсиями.
Таким образом, изменения хромосомной организации, чаще всего оказывающие неблагоприятное воздействие на жизнеспособность клетки и организма, с определенной вероятностью могут быть перс пективными, наследоваться в ряду поколений клеток и организмов и создавать предпосылки для эволюции хромосомной организации на следственного материала.
3.5.4. Значение хромосомной организации в функционировании и наследовании генетического аппарата
Хромосомный уровень организации наследственного материала обеспечивает в эукариотической клетке не только определенный ха рактер функционирования отдельных генов, тип их наследования, но и регуляцию их активности.
Хромосома как комплекс генов представляет собой эволюционно сложившуюся структуру, свойственную всем особям данного вида. Взаимное расположение генов в составе хромосомы играет немаловаж ную роль в характере их функционирования. Расположение гена в той или иной хромосоме определяет тип наследования соответствующего признака (см. разд. 6.3.1).
Принадлежность генов к одной хромосоме обусловливает сцеплен ный характер наследования детерминируемых ими признаков, а рас стояние между генами влияет на частоту рекомбинации этих признаков в потомстве (правило Т. Моргана). Расположение генов в разных
133
132

хромосомах служит основой независимого наследования признаков (закон независимого наследования признаков Г. Менделя).
Образуя в хромосоме устойчивый комплекс с гистонами, ДНК эукариотической клетки оказывается недоступной для других белков, осуществляющих транскрипцию (РНК-полимераза) и выполняющих регуляторные функции (см. разд. 3.6.6.4). У прокариот кольцевая
Междисковый |
Диск |
Рис, 3.65. Пуфы в политенных хромо |
участок |
сомах. А —схема образования пуфа; |
|
|
|
|
|
|
Б—электронограмма политенных |
|
|
хромосом слюнных желез насекомых |
|
|
Стрелками обозначены места пуфов |
РНК-транскрипты
Рис. 3.66. Хромосомы типа ламповых щеток. А — схема строения; Б — хромосомы из овоцита амфибий
молекула ДНК, формирующая неустойчивый комплекс с негистоновыми белками, остается доступной для белков, активирующих отдель ные гены путем прямого взаимодействия с определенными нуклеотидными последовательностями ДНК. Таким образом, гистоны, участвующие в пространственной организации ДНК в хромосоме, регулируют генную активность, угнетая ее.
Процесс транскрипции с участка ДНК эукариотической клетки предполагает необходимость предварительной декомпактизации хро-
134 |
135 |

Рис. 3.66. Продолжение
матина путем временного ослабления связи ДНК с этими белками. Примером такой декомпактизации хроматина являются пуфы (взду тия), наблюдаемые в участках политенных хромосом и соответствую щие активно функционирующим генам (рис. 3.65) или обнаруженные в овоцитах амфибий хромосомы типа ламповых щеток, в которых активные участки ДНК формируют петлеобразные структуры с мень шей степенью компактизации (рис. 3.66).
3.5.5. Биологическое значение хромосомного уровня организации наследственного материала
Возникновение хромосомной организации наследственного мате риала в эукариотической клетке тесно связано с большим его объемом по сравнению с прокариотической клеткой. Распределение основной массы генетического материала в ограниченном количестве ядерных структур — хромосом — обеспечивает упорядоченность его простран ственной организации по группам сцепления. Относительная самосто ятельность хромосом в процессах репликации ДНК и распределения ее молекул между дочерними клетками позволяет закономерно восп роизводить и передавать обширную информацию в ряду клеточных поколений, сохраняя постоянство ее организации. Наконец, распре деление генов по группам сцепления допускает возможность реком бинации генетического материала гомологичных и негомологичных хромосом при мейозе и оплодотворении (см. разд. 3.6.2.3).
Эффективность рекомбинации наследственного материала, возраста ющая у эукариот благодаря его хромосомной организации, существенно увеличивает степень комбинативной изменчивости у данных организмов. Это является важным эволюционным фактором, обеспечивающим раз нообразный исходный материал для естественного отбора.
3.6. ГЕНОМНЫЙ УРОВЕНЬ ОРГАНИЗАЦИИ НАСЛЕДСТВЕННОГО МАТЕРИАЛА
3.6.1. Геном. Генотип. Кариотип
Геномом называют всю совокупность наследственного материала, заключенного в гаплоидном наборе хромосом клеток данного вида организмов. Геном видоспецифичен, так как представляет собой тот необходимый набор генов, который обеспечивает формирование ви довых характеристик организмов в ходе их нормального онтогенеза. Например, у некоторых видов появляются гаплоидные организмы, которые развиваются на осно ве одинарного набора генов, заключенного в геноме. Так, у ряда видов членистоногих гап лоидными являются самцы, развивающиеся из неоплодотворенных яйцеклеток.
При половом размножении в процессе оплодотворения объединяются геномы двух ро дительских половых клеток, образуя генотип нового орга низма. Все соматические клет ки такого организма обладают двойным набором генов, полу ченных от обоих родителей в виде определенных аллелей. Таким образом, генотип — это генетическая конституция ор ганизма, представляющая собой совокупность всех на следственных задатков его клеток, заключенных в их хромосомном наборе —- кариотипе.
Кариотип —диплоидный набор хромосом, свойствен ный соматическим клеткам ор ганизмов данного вида, являющийся видоспецифическим признаком и характери зующийся определенным
л*
?. •
X,
//#
Рис. 3.67. Кариотипы организмов различных видов:
I — скерды, //—дрозофилы, ///—человека
136 |
137 |
|

числом, строением и генетическим составом хромосом (рис. 3.67). Ниже приведены количества хромосом соматических клеток некото рых видов организмов.
Если число хромосом в гаплоидном наборе половых клеток обоз начить п, то общая формула кариотипа будет выглядеть как 2п, где значение п различно у разных видов. Являясь видовой характеристикой организмов, кариотип может отличаться у отдельных особей некото рыми частными особенностями. Например, у представителей разного пола, имеются в основном одинаковые пары хромосом {аутосомы), но их кариотипы отличаются по одной паре хромосом (гетерохромосомы, или половые хромосомы). Иногда эти различия состоят в разном коли честве гетерохромосом у самок и самцов (XX или ХО). Чаще различия касаются строения половых хромосом, обозначаемых разными бук вами — X и Y (XX или XY).
Каждый вид хромосом в кариотипе, содержащий определенный комплекс генов, представлен двумя гомологами, унаследованными от родителей с их половыми клетками. Двойной набор генов, заключен ный в кариотипе,— генотип — это уникальное сочетание парных ал лелей генома. В генотипе содержится программа развития конкретной особи.
Животные |
|
Число хромосом |
Малярийный плазмодий .' |
|
2 |
Гидра |
. |
32 |
Таракан |
|
48 |
Комнатная муха" |
|
12 |
Сазан |
|
104 |
Окунь |
'.. |
28 |
Зеленая лягушка |
|
26 |
Голубь . |
|
80 |
* Кролик |
|
44 |
Шимпанзе |
|
48 |
Человек |
|
46 |
3.6.2. Проявление свойств наследственного материала
на геномном уровне его организации
3.6.2.1. Самовоспроизведение и поддержание постоянства кариотипа в ряду поколений клеток
В основе самовоспроизведения кариотипа в ряду клеточных поко лений лежат процессы самоудвоения отдельных хромосом, описанные в разд. 3.5.3.1. Жизнедеятельность клеток как структурно-функцио нальных единиц живого обеспечивается всей совокупностью получа емого ими генетического^материала, поэтому особое значение приобретает поддержание постоянства кариотипа в ряду их поколений. На геномном уровне организации наследственного материала эта обеспечивается всей совокупностью процессов, происходящих в митотическом цикле (см. гл. 2).
Если содержание ДНК в гаплоидном набо ре хромосом, т.е. в геноме, является видоспецифическим и соответствует с, то в диплоидном кариотипе, равном 2л, оно состав ляет 2с. В ходе митотического цикла количе ство ДНК в клетке изменяется от 2с (сразу после ее образования, когда хромосомы состо ят из одной нити) до 4с (после репликации ДНК в S-периоде интерфазы, когда хромосо мы становятся двунитчатыми). Число хромо сом при этом остается неизменным и равным In на протяжении всего митотического цикла. Благодаря описанным ранее процессам, про исходящим в ходе интерфазы и последующего митоза (см. гл. 2), дочерние клетки получают
от материнской полноценную наследственную программу — генотип, заключенный в кариотипе.
Однако не всегда клетки, удвоившие свой наследственный мате риал, вступают в митоз. Этим обусловлено существование в тканях некоторого количества клеток с формулой наследственного материала 2п4с, представляющих определенный резерв, готовый немедленно приступить к делению. Наконец, не всегда начавшееся деление закан чивается образованием двух дочерних клеток. Нарушение деления
Рис. 3.69. Образование полиплоидных клеток в результате разрушения веретена деления. А —различные наборы хромосом (12, 24, 48) в пыльцевых зернах одного из сортов гиацинта; Б —образование полиплоидных клеток:
/ —в норме, //—при разрушении веретена деления колхицином
138 |
139 |

цитоплазмы при нормальном формировании дочерних ядер приводит к образованию двуядерных клеток (рис. 3.68). Разрушение ахроматинового веретена, делающее невозможным распределение дочерних хромосом между его полюсами, сопровождается появлением полипло идных клеток (рис. 3.69), в которых An дочерних хромосом остаются в одном неразделившемся ядре.
Указанные отклонения от обычного течения митотического цикла являются причиной появления в тканях клеток, несущих больший по объему наследственный материал.
3.6.2.2.Механизмы поддержания постоянства кариотипа
вряду поколений организмов
Уорганизмов, размножающихся бесполым путем, новое поколение появляется из неспециализированных в отношении генеративной фун кции клеток тела. В основе их самовоспроизведения лежит митоз,
Рис. 3.70. Сравнение первого мейотического (редукционного) деления с мито зом
Профаза—спирализация хромосом, на чало формирования веретена деления; в мей озе, кроме того, происходит конъюгация гомологичных хромосом с образованием би валентов;
метафаза — в митозе в экваториальной плоскости веретена деления располагаются отдельные хромосомы числом 2л, в мейозе в плоскости экватора выстраивается л бивален тов;
анафаза —в митозе в результате расщеп ления центромер дочерние хромосомы (быв шие сестринские хроматиды) расходятся к разным полюсам (по 2л к каждому полюсу), в мейозе разрушаются биваленты и гомологи расходятся к разным полюсам (по одному из каждой пары); формируется гаплоидный на бор хромосом;
телофаза—в митозе формируются ядра дочерних клеток, в мейозе телофаза сокраще на во времени, так как не происходит полной деспирализации хромосом и клетки сразу пе реходят ко второму делению.
Результаты митоза — сохранение в дочер них клетках диплоидного набора хромосом (2л2с); результаты первого мейотического де ления— образование клеток с гаплоидным •набором двунитчатых хромосом (nlc)
обеспечивающий таким образом сохране ние постоянной структуры наследствен ного материала в ряду поколений не толь ко клеток, но и организмов.
При половом размножении процесс воспроизведения организмов осуществ ляется с участием специализированных половых клеток — гамет, вступающих в оплодотворение. При оплодотворении на следственный материал двух родитель ских гамет сливается, образуя генотип организма нового поколения — зиготы. Чтобы потомки получили соответствую щую программу для развития видовых и индивидуальных характеристик, они дол
жны обладать кариотипом, которым располагало предыдущее поколе ние. В такой ситуации поддержание постоянства кариотипа в ряду поколений организмов достигается предварительным уменьшением вдвое набора хромосом в гаметах, который восстанавливается до диплоидного при их оплодотворении: п + п = 2л.
Образование гаплоидных гамет осуществляется в ходе гаметогенеза путем особой формы клеточного деления —мейоза. При мейозе из клеток с диплоидным набором 2л образуются гаметы с гаплоидным набором хромосом п (см. гл. 5). Такой результат достигается благодаря тому, что после однократного удвоения ДНК клетка делится дважды. В отличие от митоза в первом мейотическом делении в результате конъюгации гомологичные хромосомы объединяются в пары — бива ленты. Последующее расхождение гомологов к разным полюсам вере тена деления приводит к образованию клеток с гаплоидным набором хромосом: 2пАс -> «2с. На рис. 3.70 представлены особенности первого деления мейоза в сравнении с митозом. В ходе второго мейотического деления сестринские хроматиды каждой хромосомы, как и в митозе, распределяются между дочерними клетками с наследственным мате риалом пс (рис. 3.71).
Благодаря особенностям мейоза образуются клетки, несущие пол ноценный геном, в котором каждая группа сцепления представлена в единственном экземпляре (гаплоидный набор хромосом).
При самооплодотворении гаметы одного и того же родителя, а при перекрестном оплодотворении половые клетки разных организмов взаимодействуют друг с другом. Сперматозоиды, проникая в яйцек летку, вводят в нее свой ядерный наследственный материал, заклю ченный в гаплоидном наборе хромосом. Ядра гамет сливаются и формируют диплоидное ядро зиготы, в котором каждая группа сцеп ления представлена в двойном экземпляре —отцовской и материн ской хромосомами.
Таким образом, мейоз и последующее оплодотворение обеспечи вают сохранение у нового поколения организмов диплоидного кари отипа, присущего всем особям данного вида.
140 |
141 |