

61. Биосинтез триглицеридов
Синтез триглицеридов в тканях происходит с учетом двух путей образования глицерол-3-фосфата и возможности синтеза триглицеридов в стенке тонкой кишки из в-моноглицеридов, поступающих из полости кишечника в больших количествах после расщепления пищевых жиров. Существуют глицерофосфатный, дигидроксиацетонфосфатный и в-моноглицеридный пути синтеза триглицеридов. Большинство ферентов, участвующих в биосинтезе ТГ, находятся в эндоплазматическом ретикулуме, но некоторые, например глицерол-3-фосфатацилтрансфераза, находятся в митохондриях.
62. Биосинтез ВЖК протекает в ЭПС клеток. Исходным материалом для синтеза заменимых ВЖК является АЦЁТИЛ-КОА.
Условия для биосинтеза ВЖК:
1.Наличие АТФ, СО2, Н2О, НАДФ*Н2, поступающий из ГЕКСОЗОМОНОФОСФАТНОГО пути превращения глюкозы.
2.Наличие специальных белков-переносчиков (HS-АПБ).
3.Наличие специальных ферментов синтеза.
Процесс биосинтеза циклический. Каждый цикл включает в себя 6 этапов. АЦЕТИЛ-КОА используется на первом этапе, как «затравка» синтеза.
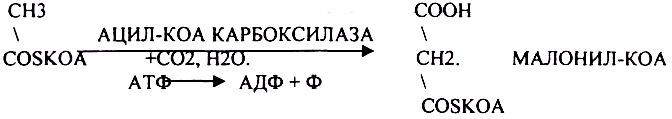
Первый этап. Этап образования 3-углеродного соединения - МАЛОНИЛ-КОА.
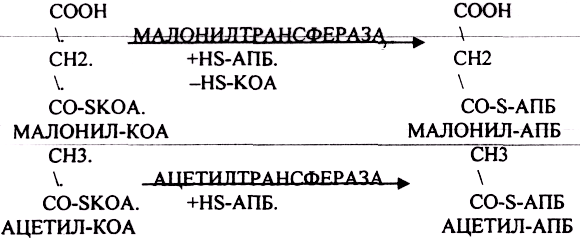
Второй этап. Этап переноса МАЛОНИЛА и АЦЕТИЛА на специальные белки - HS-АПБ.
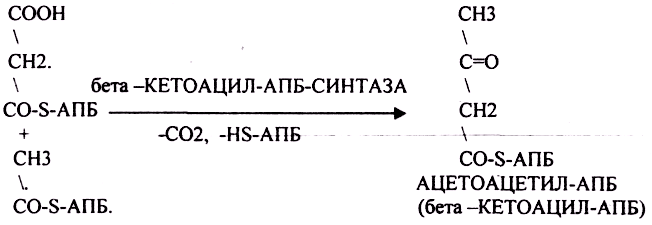
Третий этап. Этап конденсации МАЛОНИЛА-АПБ и АЦЕТИЛА-АПБ с участием ТРАНСФЕР АЗЫ - СИНТАЗЫ.
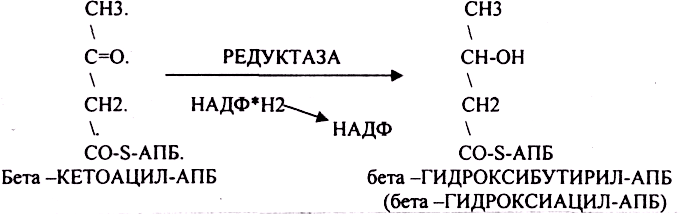
Четвёртый этап. Этап восстановления бета -КЕТОАЦИЛ-АПБ.
Пятый этап. Этап ДЕГИДРАТАЦИИ бета-ГИДРОКСИАЦИЛ-АПБ.

Шестой этап. Этап восстановления ЕНОИЛАЦИЛ-АПБ.
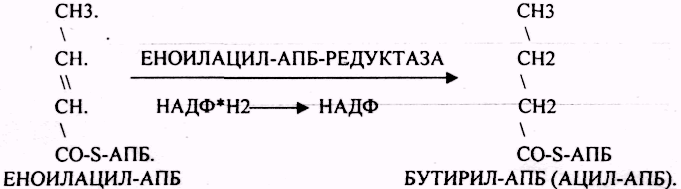
 Т.о.
завершается 1 цикл синтеза ВЖК образованием
масляной кислоты (БУТИРИЛ-АПБ). В
дальнейшем последовательно и циклично
к 4-углеродному фрагменту будут
присоединяться молекулы МАЛОНИЛ-КОА.
Для подсчёта количества циклов синтеза
ВЖК, подсчёта молекул АТФ, МАЛОНИЛ-КОА
можно пользоваться формулой:N/2
- 1, гдеN-это число углеродных
звеньев в молекуле ВЖК. Завершается
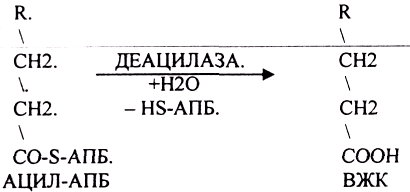
биосинтез любой ВЖК в тканях ДЕАЦИЛАЗНОЙ
реакцией.
Т.о.
завершается 1 цикл синтеза ВЖК образованием
масляной кислоты (БУТИРИЛ-АПБ). В
дальнейшем последовательно и циклично
к 4-углеродному фрагменту будут
присоединяться молекулы МАЛОНИЛ-КОА.
Для подсчёта количества циклов синтеза
ВЖК, подсчёта молекул АТФ, МАЛОНИЛ-КОА
можно пользоваться формулой:N/2
- 1, гдеN-это число углеродных
звеньев в молекуле ВЖК. Завершается
биосинтез любой ВЖК в тканях ДЕАЦИЛАЗНОЙ
реакцией.
Так в организме человека происходит синтез всех заменимых ВЖК - всех предельных и непредельных ВЖК, имеющих одну двойную связь.
63. 24Интеграция углеводного и липидного обмена
В любой живой системе обмен липидов связан с обменом соединений других классов,например, обменом углеводов или аминокислот. Эти взаимосвязи можно проследить, воспользовавшись общей схемой обмена липидов:
┌─────── Пищевые липиды ───────┐
│ │ │
│ │ │
┌─────│───> Высшие жирные кислоты <──│────────┐
Структурные │ │ │ Резервные
липиды │ │ │ липиды
^ ^ │ │ ^ ^
│ └─────┼───────── Ацил-КоА ───────────┼────────┘ │
Глицерол │ │ ^ │ │
Аминоспирты <──┘ │ │ └───>Глицерол
^ │ │ ^
├─────────────────> Ацетил-КоА <────────────────┤
│ ││ │ │ │
Глюкоза ││ │ │ Ацетоновые Глюкоза
Аминокислоты СО + Н <─┘│ │ └─> тела Аминокислоты
│ │
Стероиды <────┘ └─────> Полипреноиды
Из схемы следует, что пищевые липиды являются источниками высших жирных кислот, глицерола, аминоспиртов и некоторых других соединений, используемых организмом для синтеза свойственных для
него структурных или резервных липидов. Свободные высшие жирные кислоты, наряду с глицеролом и аминоспиртами образуются в организме также при расщеплении резервных или структурных липидов.
Еще одним источником высших жирных кислот может служить их синтез из ацетила-КоА, который в свою очередь может быть промежуточным продуктом обмена углеводов или аминокислот. Заметим, что жирные кислоты в клетках используются в различных метаболических путях клетки только в их активированной форме - в виде ацил-КоА.
Одним из ключевых метаболитов липидного обмена является ацетил-КоА, поскольку, во-первых, именно через это соединение осуществляется окислительное расщепление высших жирных кислот; во-вторых, через ацетил-КоА атомы углерода жирных кислот могут быть использованы для пластических целей - для синтеза холестерола или полипреноидов; в третьих, через ацетил-КоА в гепатоцитах углеродные цепи жирных кислот преобразуются в ацетоновые тела гидрофильные "топливные" молекулы, легко транспортируемые в клетки различных органов и тканей; в четвертых, через ацетил-КоА осуществляются метаболические превращения углеродных скелетов аминокислот и моносахаридов в жирные кислоты, используемые в дальнейшем для синтеза более сложных липидных молекул.
Соединения других классов - аминокислоты и моносахариды – в ходе своего метаболизма образуют промежуточные продукты, которые могут в дальнейшем использоваться в клетке как для синтеза высших
жирных кислот, так и для образования других мономерных единиц, необходимых для синтеза сложных липидов: глицерола, этаноламина, холина, сфингозина и пр. Таким образом, обмен липидов оказывается тесно связанным с обменом соединений других классов, а метаболические пути обмена липидов различных классов являются частью общей метаболической сети, функционирующей в организме.
Известно, что в постабсорбционном состоянии основным энергетическим "топливом" для клеток являются высшие жирные кислоты, тогда как в период пищеварения, когда во внутреннюю среду организма поступают моносахариды и ресинтезированные в стенке кишечника триглицериды, основным энергетическим топливом становится глюкоза; более того, поступающая в клетки глюкоза превращается в жирные кислоты. Последний процесс особенно характерен для гепатоцитов и липоцитов.
64. 252жиро-углеводный цикл рэндла. Цикл глюкоза — свободные жирные кислоты — цикл Рэндла — является одним из механизмов, обеспечивающих гомеостаз глюкозы. Повышенное окисление свободных жирных кислот в мышечной ткани приводит к истощению запасов НФД+, что способствует ингибированию цикла Кребса и увеличению внутриклеточного содержания цитрата и ацетил-СоА. Последний в свою очередь угнетает активность пируватдегидрогеназы, вследствие чего пируват конвертируется в лактат или аланин. Повышенное содержание цитрата, которое образуется в цикле Кребса из ацетил-СоА и оксалоацетата, угнетает активность фосфофруктокиназы, способствуя повышению концентрации глюкозо-6-фосфата, ингибируя гексокиназу, снижая утилизацию глюкозы и соответственно гликолиз.
65. Механизм всасывания, активации и мембранного транспотрта ЖК в митохондрии.В постабсорбционном состоянии основным энергетическим "топливом" для клеток являются высшие ЖК (поступающая в клетки глюкоза превращается в жирные кислоты). При поступлении глюкозы в клетки она в цитозоле окисляется до пирувата ,последний окисляется в матриксе митохондрий до ацетил-КоА. Образовавшийся ацетил-КоА конденсируется с оксалоацетатом (ЩУК) с образованием цитрата, а цитрат выходит из митохондрии в цитозоль. Поступивший в цитозоль цитрат, во-первых, служит источником ацетил-КоА и восстановительных эквивалентов для синтеза высших жирных кислот, а, во-вторых, активирует фермент ацетил-КоА-карбоксилазу, стимулируя тем самым образование малонил-КоА, также необходимого для синтеза высших жирных кислот. В результате при избытке глюкозы в клетке запускается синтез жирных кислот. Малонил-КоА в свою очередь угнетает перенос высших жирных кислот из цитозоля в матрикс митохондрий, ингибируя активность внешней ацетил-КоА: карнитин-ацилтрансферазы, выключая таким образом окисление высших жирных кислот. В итоге при поступлении глюкозы в клетку угнетается окисление высших жирных кислот, стимулируется их синтез, а потребность клетки в энергии покрывается за счет аэробного окисления глюкозы, чему способствует повышение концентрации ацетил-КоА и цитрата в матриксе митохондрий. Увеличение концентрации жирных кислот в клетке наряду с нарастанием концентрации в них триозофосфатав создает условия для синтеза резервных триглециридов. В этот процесс включаются также высшие жирные кислоты и глицерол, поступающие в клетку в результате гидролиза триглицеридов ХМ и ЛПОНП. Основная роль карнитина – активное расщепление жиров и обеспечение организма необходимой энергией: L-карнитин доставляет жирные кислоты в митохондрии клеток, где они достаточно быстро подвергаются окислительной реакции.
Гиперхолестеринемия.Гиперхолестеринемия (ГПЕ) – повышенный уровень холестерина в крови. Гиперхолестеринемия относится к первичным нарушениям липидного обмена. Пример заболевания ГПЕ: семейная гиперхолестеринемия- при этом заболевании в организме нарушен синтез рецепторов для ЛПНП, в результате чего нарушена утилизация этих липопротеидов. В крови таких больных повышено содержание ЛПНП и холестерола. Тяжесть заболевания в значительной мере зависит от того, один или оба гена белков-рецепторов ЛПНП дефектны. При дефекте одного из генов в клетках имеется половинное количество рецепторов для ЛПНП, если дефектны оба гена - рецепторов для ЛПНП вообще нет. Виды ГПЕ: а) неосложненный ( физическое старение, старость, смерть)- накопление ХС в плазматических мембранах, как результат снижения стероидогенеза ( секс-гормонов); б) осложненный (атеросклероз ) в форме ИБС ( инфаркт миокарда, кардиосклероз), ишемии мозга ( инсульт, тромбоз), ишемии конечностей ( гангрена), ишемии органов и тканей, дегенерации брадитрофных структур ( катаракта, остеохондроз), связанный с уменьшением желчегенеза. Атеросклероз — это патологическое состояние, при котором происходит отложение липидов в стенке сосудов средних и больших размеров. Коэффициент атерогенности (Ка) - расчетный показатель степени риска развития атеросклероза у человека. Определяется по количеству в крови липопротеидов низкой плотности (ЛПНП), липопротеидов высокой плотности (ЛПВП), общего холестерина. В норме показатель менее 3-х. Ка = холестерин ЛПНП + холестерин ЛПОНП/холестерин ЛПВП.
Липидозы (липоидозы) - наследственные заболевания нервной системы, обусловленные расстройством обмена липидов. В основе патогенеза липидозов - генетически детерминированные разнообразные ферментативные дефекты, которые приводят к тяжелым нарушениям в организме. Условно эти заболевания подразделяются на болезни накопления (внутриклеточные липидозы, при которых преимущественно поражаются нервные клетки, и вторично - проводящие пути) и лейкодистрофии (прогрессирующий распад белого вещества). В основе Л. лежит полная или частичная недостаточность лизосомальных ферментов, участвующих в обмене липидов и обусловленная наследственным дефектом соответствующего гена. Большинство Л. наследуется по аутосомно-рецессивному типу, исключение составляет болезнь Фабри, которая наследуется по Х-сцепленному, рецессивному типу. К болезням накопления (внутриклеточным липидозам) относятся амавротичес-кая идиотия, болезнь Ниманна-Пика, болезнь Гоше и болезнь Рефсума.
Патология липидного обмена возникает по двум причинам:а) при нарушении переваривания и всасывания липидов;б) при нарушении метаболизма липидов и липопротеидов.
В основе нарушения переваривания и всасывания липидов лежат три группы патологических процессов:1) в поджелудочной железе, сопровождающиеся дефицитом панкреатической липазы; 2) в печени- при закупорке желчных протоков и фистуле желчного пузыря, приводящие к дефициту желчи; 3) в кишечнике, сопровождающиеся снижением метаболической активности слизистой оболочки, где локализованы ферменты синтеза ТГ. В соответствии с этим различают панкреатическую, гепатогенную и энтерогенные формы стеаторей. Стеаторея- нарушение переваривания жиров. Панкреатическая стеаторея вызывается дефицитом панкреатической липазы, что наблюдается при хроническом панкреатите, врожденной гипоплазии пакреас и муковисцедозе, когда наряду с другими железами поражена и поджелудочная железа.
Гепатогенная стеаторея наблюдается при врожденной атрезии желчных путей, механических желтухах, гепатитах, циррозе.
Энтерогенная стеаторея отмечается при целиакии, абеталипопротеинемии, интестинальной лимфангиэктазии, интестинальной липодистрофии, амилоидозе и обширной резекции тонкого кишечника.
При переваривании пищевых жиров в пищеварительном тракте высвобождаются моноглицериды и высшие жирные карбоновые кислоты (ВЖК)., которые после всасывания образуют ТГ. Последние являются основными компонентом хиломикронов (ХМ). Липиды, являясь гидрофобными , транспортируются кровью в виде особых надмолекулярных образований- липопротеидов(ЛП), в состав которых входят ХС, ТГ, ФЛ (фосфолипиды) и апобелки.
Диагностическое значение имеет определение в крови содержания ТГ, СЖК, ХС, желчных кислот, ФЛ и ЛП а также состав последних.
Ожирение.
Это состояние характерно, прежде всего, для жителей развитых стран, у которых на 100% выше вероятность преждевременной (раньше, чем средняя продолжительность жизни) смерти.
У людей сизбыточным весом эта вероятность выше на 10% - 25%.
С проблемой ожирения тесно связаны вопросы гуморальной регуляции синтеза и распада ТГ в жировой ткани.
Ожирение развивается тогда, когда поступление в организм энергосубстратов превышает расходование энергии. Среди причин развития этого состояния, включающих генетические и средовые факторы,
Важнейшее значение имеют состав принимаемой пищи и специфические регуляторы энергетического баланса. Одним из таких соединений являетсяпродукт экспрессии так называемого гена ожирения(ob - гена) - лептин (leptos – греч. «тонкий»).
Лептин является белком, состоящим из 167 аминокислот. Местом синтезалептина являются адипоциты, в гораздо меньшей степени - эпителиальные клетки желудка и плаценты. Причем, чем больше в этих клетках накапливается ТГ, тем больше там образуетсялептина..
Действие лептина на депонирование ТГ в жировой ткани опосредовано его влиянием на гипоталамические центры, контролирующие поведение и чувство голода, температуру тела и энергозатраты