


Распад пуринов и перекисные процессы
Генерация активных форм кислорода при ишемии- реперфузии

Ишемия. Распад АТФ до гипоксантина
• прекращение кровотока (ишемия) сопровождается гипоксией;
• распад АТФ начинает преобладать над его синтезом:
АТФ
АДФ |
|
|
|
|
NH2 |
|
N |
|
|
|
|
|
|
|
|
|
|
|
|
O |
|
|
|
N |
|
|
|
|
|
|
|
|
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
N |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||
|
O |
|
|
|
|
|
|
|
|
|
|
HN |
|
|
|
|
|
O |
|
||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
N |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
N |
|
|
|
|
|||||||||||
|
|
|
|
N |
|
|
O |
|
|
|
|
|
N |
|
|
|
N |
||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
-O |
|
P |
|
O |
|
O |
|
|
|
|
-O |
|
|
P |
|
O |
|
|
|
|
|
|
|
O |
|
|
|
|
|
HN |
|
|
|
|
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
O- |
|
|
H |
H |
|
|
O- |
|
|
|
|
H |
|
|
H |
|
|
|
|
|
|
|
|
N |
||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
H |
|
|
|
|
|
|
|
H |
|
|
|
|
|
|
|
|
|
H |
|
|
|
|
|
|
|
H |
|
|
|
N |
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
OH |
|
|
|
|
|||||||||||||||||||
|
|
|
|
|
|
|
OH |
OH |
|
|
|
|
|
|
|
|
|
|
|
OH |
|
|
|
H |
|||||||||||||||||||
|
|
|
|
АМФ |
аденилатдезаминаза |
ИМФ |
нуклеозидфосфорилаза |
|
|||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Гипоксантин |
|||||||||||||||||||||||||
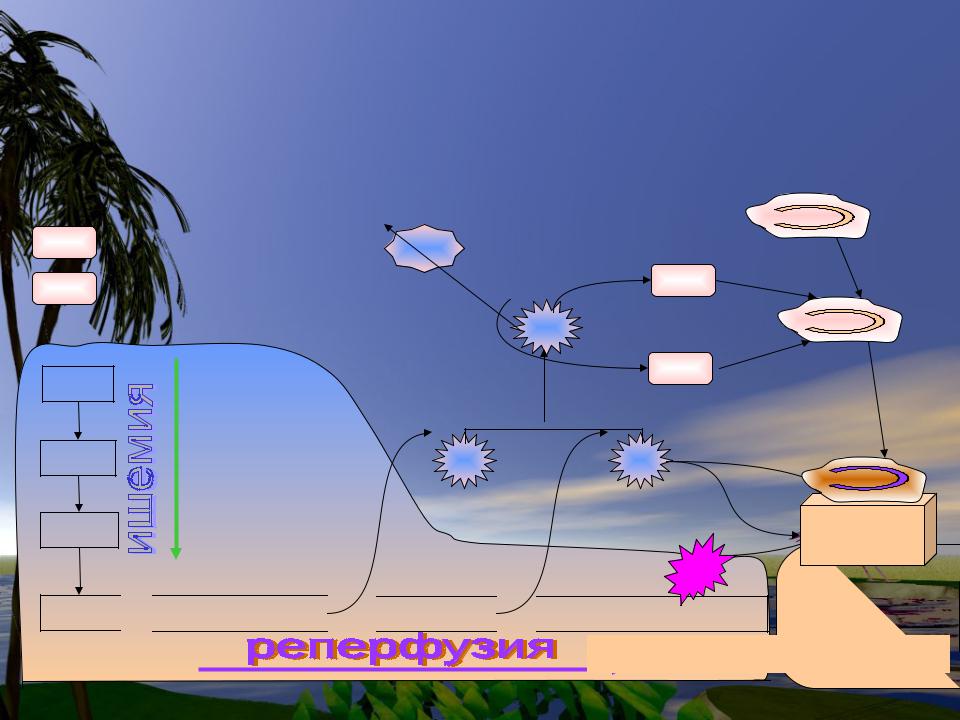
Повреждение тканей при ишемии- реперфузии
|
|
|
Нейтрофил |
|
OH |
- гидроксильный радикал H2O |
Fe2+ |
|
|
HOCl |
- гипохлорит |
OH |
||
|
||||
|
|
|||
|
Каталаза H2O2 |
|
||
АТФ |
СОД |
|
HOCl |
|
Миелопероксидаза |
||||
|
||||
АДФ |
O2 - |
|
O2 - |
|
АМФ |
|
|
CD11/CD18 |
|
|
|
—————— |
||
|
|
|
ELAM |
|
ИМФ 
 Гипоксантин
Гипоксантин 
 Ксантин
Ксантин 
 Мочевая кислота
Мочевая кислота
Эндотелиальные клетки

Реперфузия. Окисление гипоксантина до мочевой кислоты
•После восстановления кровотока (реперфузия) в ткани начинает поступать кислород;
•окисление гипоксантина до мочевой кислоты сопровождается образованием активных форм кислорода:
O |
|
H2O+O2 H2O2 |
O |
|
H2O+O2 |
H O |
2 |
O |
H |
|
|
|
|
N |
|
2 |
|
||
HN |
N |
|
HN |
|
|
HN |
|
N |
|
|
|
|
|
|
O |
||||
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
N |
N |
ксантиноксидаза |
O N |
N |
|
|
O |
N |
N |
H |
H |
H |
ксантиноксидаза |
H |
H |
||||
Гипоксантин |
Ксантин |
Мочевая кислота |

Биосинтез
пиримидиновых
нуклеотидов

•Непосредственным предшественником при синтезе пиримидиновых колец является карбомоилфосфат, который
образуется из Глу и НСО3- и аспартата.


Реакции 1,2,3 протекают в цитоплазме и катализируются одним полифункциональным ферментом ( имеющим 3 АЦ). На 4- стадии дигидрооротат окисляется FMN- зависимой дегидрогеназой в оротат,
который связывается с ФРПФ (FRPF), образуя оротидиловую кислоту.

Последняя декарбоксилируется тем же ферментом, образуя УМФ.
Т.о. 6 катализирующих активных центров кодируются только тремя структурными генами.