

Строение фоторецепторов
Основными светочувствительными элементами (рецепторами) являются два вида клеток: одни в виде стебелька - палочки 110-123 млн. (высота 30 мкм, толщина 2мкм), другие более короткие и более толстые - колбочки 6-7 млн. (высота 10мкм, толщина 6-7 мкм). Они распределены в сетчатке неравномерно. Центральная ямка сетчатки( fovea centralis) содержит только колбочки(до 140 тыс. на 1 мм 52 0). По направлению к периферии сетчатки их число уменьшается, а число палочек возрастает.
Каждый фоторецептор - палочка или колбочка состоит из чувствительного к действию света наружного сегмента содержащего зрительный пигмент и внутреннего сегмента, который содержит ядро и митохондрии обеспечивающие энергетические процессы в фоторецепторной клетке
Колбочки функционируют в условиях больших освещенностей - обеспечивают дневное и цветовое зрение, а палочки - отвечают за сумеречное зрение .

Видимый нами спектр электромагнитных излучений заключен между коротковолновым (длина волны от 400нм) излучением, которое мы называем фиолетовым цветом и длинноволновым излучением(длина волны до 700 нм) называемым красным цветом. В палочках находится особый пигмент- родопсин , (состоит из альдегида витамина А или ретиналя ибелка) или зрительный пурпур, максимум спектра, поглощения которого находится в области 500 нанометров. Он ресинтезируется в темноте и выцветает на свету. При недостатке витамина А нарушается сумеречное зрение -"куриная слепота".
В наружных сегментах трех типов колбочек(сине-, зелено и красно-чувствительных) содержится три типа зрительных пигментов, максимум спектров поглощения которых находится в синей (420 нм), зеленой(531 нм) и красной(558 нм) частях спектра. Красный колбочковый пигмент получил название - " йодопсин" . Структура йодонсина близка к родонсину.
Молекулярная физиология фоторецепции: Внутриклеточные регистрации от колбочек и палочек животных показали, что в темноте вдоль фоторецептора течет темновой ток, выходящий из внутреннего сегмента и входящий в наружный сегмент. Освещение приводит к блокаде этого тока. Рецепторный потенциал модулирует выделение медиатора (глутамата) в синапсе фоторецептора. Было показано, что в темноте фоторецептор непрерывно выделяет медиатор, который действует деполяризующим образом на мембраны постсинаптических отростков горизонтальных и биполярных клеток.(см. Рис.1)
Рис.1
Рис 2.

Палочки и колбочки обладают уникальной среди всех рецепторов электрической активностью, их рецепторные потенциалы при действии света - гиперполяризующие, потенциалы действия под их влиянием не возникают.
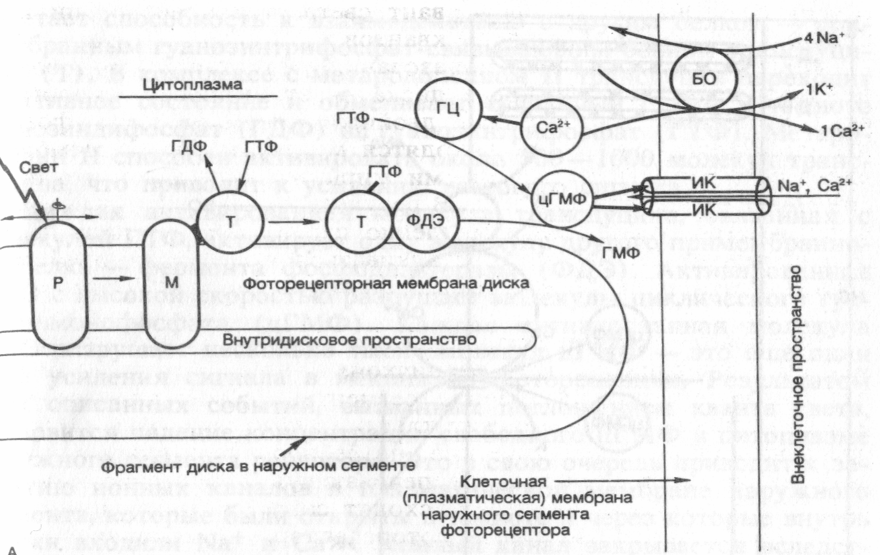
{ При поглощении света молекулой зрительного пигмента.(см. Рис.2) - родопсина в ней происходит мгновенная изомеризация ее хромофорной группы: 11- цис -ретиналь превращается в транс -ретиналь. Вслед за за фотоизомеризацией ретиналя происходят пространственные изменения в белковой части молекулы: она обесцвечивается и переходит в состояние метородопсина II. В результате этого молекула зрительного пигмента приобретает способность к взаимодействию с другим примембранным белком гуанозин трифосфат(ГТФ) - связывающим белком – трансдуцином (Т).
В комплексе с метародопсином трансдуцин переходит в активное состояние и обменивает связанный с ним в темноте ганозитдифосфат(ГДФ) на (ГТФ). Трансфдуцин + ГТФ, активируют молекулу другого примеммбранного белка - фермента фосфодиэстеразы(ФДЭ). Активированная ФДЭ разрушает несколько тысяч молекул цГМФ.
В результате падает концентрация цГМФ в цитоплазме наружного сегмента рецептора. Это приводит к закрытию ионных каналов в плазматической мембране наружного сегмента, которые были открыты в темноте и через которые внутрь клетки входили Na + и Ca . Ионные каналы закрываются вследствие того, что падает концентрация цГМФ, которая держала каналы открытыми. В настоящее время выяснено, что поры в рецепторе открываются благодаря цГМФ циклическому гуанозинмонофосфату.
Механизм восстановления исходного темного состояния фоторецептора связан с повышением концентрации цГМФ. (в темновую фазу с участием алкагольдегидрогеназы + НАДФ)
Т.о поглощение света, молекулами фотопигмента приводит к снижению проницаемости для Nа, что сопровождается гиперполяризацией, т.е. возникновением рецепторного потенциала. Гиперполяризационный рецепторный потенциал, возникший на мембране наружного сегмента, распространяется затем вдоль клетки до ее пресинаптического окончания и приводит к уменьшению скорости выделения медиатора - глутамата. Кроме глутамата нейроны сетчатки могут синтезировать и другие нейромедиаторы, такие как ацетилхолин, дофамин, глицин ГАМК.
Фоторецепторы связаны между собой - электрическими(щелевыми) контактами. Эта связь избирательная: палочки связаны с палочками и т.д.
Эти ответы от фоторецепторов сходятся на горизонтальные клетки, которые приводят к деполяризации в соседних колбочках возникает отрицательная обратная связь, которая повышает световой контраст.
На уровне рецепторов происходит торможение и сигнал колбочки перестает отражать число поглощенных фотонов, а несет информацию о цвете, распределении и интенсивности света, падающего на сетчатку в окрестностях рецептора.
Существует 3-и типа нейронов сетчатки - биполярные, горизонтальные и амакриновые клетки. Биполярные клетки непосредственно связывают фоторецепторы с ганглиозными клетками, т.е. осуществляют передачу информации через сетчатку в вертикальном направлении. Горизонтальные и амакриновые клетки передают информацию по горизонтали.
Биполярные клетки занимают в сетчатке стратегическую позицию, поскольку все сигналы, возникающие в рецепторах поступающие к ганглиозным клеткам, должны пройти через них.
Экспериментально было доказано, что биполярные клетки имеют рецептивные поля в которых выделяют центр и переферию (Джон Даулинг- и др. Гарвардская медицинская школа).
Рецептивное поле - совокупность рецепторов, посылающих данному нейрону сигналы через один или большее число синапсов.
Размер рецептивных полей: d=10 мкм или 0,01 мм - вне центральной ямки.
В самой ямке d=2,5мкм (благодаря этому мы способны различать 2-е точки при видимом расстоянии между ними лишь 0,5 угловых минут-2,5мкм - если сравнить, то это монета в 5 копеек на расстоянии около 150 метров)
Начиная с уровня биполярных клеток нейроны зрительной системы дифференцируются на две группы, противоположным образом реагирующие на освещение и затемнение:
1 - клетки, возбуждающиеся при освещении и тормозящиеся при затемнении "on"- нейроны и
- клетки возбуждающиеся при затемнении и тормозящиеся при освещении - " off"- нейроны. Клетка с on-центром разряжается с заметно повышенной частотой.
Если слушать разряды такой клетки через громкоговоритель, то сначала вы услышите спонтанную импульсацию, отдельные случайные щелчки, а затем после включения света, возникает залп импульсов, напоминающий пулеметную очередь. Наоборот в клетках с off-реакцией (при выключении света - залп импульсов) Такое разделение сохраняется на всех уровнях зрительной системы, до коры включительно.
В пределах самой сетчатки передача информации осуществляется безимпульсным путем (распространением и транссинаптической передачей градуальных потенциалов).
В горизонтальных, биполярных и амокриновых клетках переработка сигнала происходит путем медленных изменений мембраны потенциалов(тонический ответ). ПД не генерируется.
Ответы палочек, колбочек и горизонтальных клеток являются гиперполяризующими, а ответы биполярных клеток могут быть как гиперполяризующие, так и деполяризующие. Амакриновые клетки создают деполяризующие потенциалы.