

09-12-Углеводы-big
.pdfКафедрабиохимииГомГМУ,2012 |
08.03.2012 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
08.03.2012 |
152 |
08.03.2012 |
153 |
|
|
Пентозный цикл ( окислительная часть) |
|
НАДФ+ |
НАДФН•Н+ |
Глюкозо-6-фосфат- |
|
дегидрогеназа |
|
Глюкозо-6-фосфат |
6-фосфоглюконо- |
|
δ-лактон |
08.03.2012 |
154 |
51

КафедрабиохимииГомГМУ,2012 |
08.03.2012 |
Вторая реакция
гидролиз 6-фглюконолактона
глюконолактонгидролазой.
глюконолактоназа
+ Н2О
6-фосфоглюконолактон 6-фосфоглюконат
08.03.2012 |
155 |
СООН |
|
|
|
|
│ |
|
|
|
|
Н–С–ОН |
НАДФ+ |
НАДФ |
|
СН2ОН |
│ |
Н∙Н+ |
|
СО2 |
│ |
Н–С–ОН |
|
|
|
С=О |
│ │
│
Н–С–ОН |
6-фосфоглюконатдегидрогеназа |
Н–С–ОН |
│ |
(декарбоксилирующая) |
│ |
Н–С–ОН |
|
Н–С–ОН |
││
СН2ОРО3Н2 |
СН2ОРО3Н2 |
6-фосфоглюконат |
Рибулозо-5-фосфат |
08.03.2012 |
156 |
Неокислительная часть.
В отличие от первой, окислительной, все реакции этой части ПФП обратимы
•Рибулозо-5-ф может изомеризоваться (фермент – кетоизомераза) в рибозу- 5-ф и эпимеризоваться (фермент – эпимераза) в ксилулозо-5-ф.
•Далее следуют 2 реакции: транскетолазная и трансальдолазная.
08.03.2012 |
157 |
52

КафедрабиохимииГомГМУ,2012 |
08.03.2012 |
• В неокислительной части рибулозо-5ф превращается в различные моносахара
С3, 4, 5, 6, 7 и 8;
• конечными продуктами являются
фр-6-ф и 3-ФГА.
08.03.2012 |
158 |
•Транскетолаза (кофермент – ТПФ)
отщепляет 2С-фрагмент и переносит его на другие сахара (см. схему). В реакции переносится 2С-фрагмент от ксилулозо-5- ф на рибозо-5-ф.
•Трансальдолаза способна переносить 3Сфрагменты.
08.03.2012 |
159 |
Рибозо-5-ф Ксилулозо-5-ф
|
Транскетолаза (ТПФ) |
Седогептулозо-7-ф |
3-ФГА |
ТРАНСКЕТОЛАЗА (КОФЕРМЕНТ –ТПФ)
- ОТЩЕПЛЯЕТ 2С-ФРАГМЕНТ И ПЕРЕНОСИТ ЕГО НА ДРУГИЕ САХАРА
- В ДАННОЙ РЕАКЦИИ ПЕРЕНОСИТСЯ 2С-ФРАГМЕНТ ОТ КСИЛУЛОЗО-5-Ф НА РИБОЗО-5-Ф.
08.03.2012 |
160 |
53

КафедрабиохимииГомГМУ,2012 |
08.03.2012 |
•Затем два образовавшиеся соединения реагируют друг с другом в трансальдолазной реакции; при этом в результате переноса 3С-фрагмента от седогептулозо-7-фосфата на 3-ФГА (3- фосфоглицериновый альдегид) образуются эритрозо-4-фосфат и фруктозо-6-фосфат.
08.03.2012 |
161 |
• Однако реакция может идти и по другому пути. В этом случае в трансальдолазной реакции образуется октулозо-1,8-дифосфат.
08.03.2012 |
162 |
|
Г л ю к о зо -6 -ф о с ф ат |
|
|
|
|
Н А Д Ф + |
Г л ю к о з о- 6-ф ос ф а т д еги д ро ген а з а |
||
|
|
|||
|
Н А Д Ф Н ·Н + |
|
|
|
|
6- ф ос ф о гл ю к о н ол а к т о н |
|
|
|
|
|
Л ак т он а з а |
|
|
|
6 -ф ос ф ог л ю к о н а т |
|
|
|
|
Н А Д Ф + |
6 -ф о с ф о гл ю ко н ат де ги др о ге н а за |
||
|
|
|||
|
Н А Д Ф Н ·Н + |
С О 2 |
|
|
|
Р и б ул о з о- 5- ф ос ф а т |
Э п и м е р аз а |
|
|
|
|
И зо м ер а за |
|
|
А р а б и н оз о -5 -ф ос ф а т |
Р и б оз о -5 -ф о сф а т |
К си л ул о з о- 5- ф ос ф а т |
||
|
|
|
Т р а н ск е т ол аза |
|
Т р а н са л ьд ол а за |
3 -Ф Г А |
С ед о ге п т у л о зо -7 -ф о сф ат |
||
|
|
|||
|
|
Ф Д А |
|
|
|
О к т ул о з о- 1,8- ди ф о сф ат |
|
|
|
|
О к ту л о зо - 8-ф о сф а т |
С ед о ге п т у л о зо -1 ,7- ди ф о с ф ат |
||
|
|
|
||
|
Т ра н с ке т о л аза |
Э р и т р о з о- 4-ф о сф а т |
Ф Д А |
|
|
|
|||
Г л ю к о з о- 6- ф о сф ат |
Ф р у к т о зо -6 -ф о сф а т |
3 -Ф Г А |
||
|
||||
08.03.2012 |
|
|
|
163 |
54

КафедрабиохимииГомГМУ,2012 |
08.03.2012 |
1.Рибозо-5Ф
предшественник б/с нуклеотидов
2.Превращение Кси5Ф, рибозо-5Ф в Ф- 6Ф и 3ФГА зависит от потребности клетки в нуклеотидах
3.ПФП активен в быстроделящихся клетках (эмбр,
регенерирующие,
08.03.2012 |
164 |
опухолевые) |
|
Итоговое уравнение ПФП
6Г-6Ф + 12NADP+  5 Г-6Ф + 12NADPH+ 6 CO2
5 Г-6Ф + 12NADPH+ 6 CO2
08.03.2012 |
165 |
Биологическая роль ПЦ
• ПЦ протекает в цитоплазме и NADРH2 не проникает и окисляется в Мх,
поэтому он не имеет энергетического
значения и выполняет только
пластическую роль.
• В ПЦ образуется 50% всего NADРH2, который используется в б/с :
08.03.2012 |
166 |
55

КафедрабиохимииГомГМУ,2012 |
08.03.2012 |
NADPH+H+
•М/с окисление
•Б/с ЖК
•Б/с ХС и стероидов (гормоны, вит D)
•Б/с аминокислот, гормонов, биогенных аминов
•Реакции фагоцитоза
•АОЗ → регенерация GSH
•Восстановление metHb (Fe3+→ Fe2+)
08.03.2012 |
167 |
Пентозы (рибоза, дезоксирибоза, ксилоза и др.)
ПЦ поставляет пентозы для синтеза:
•Моно- (FMN, АМФ, АДФ, АТФ и аналогов)
•Ди (NAD, NADP, FAD) и
•Полинуклеотидов (ДНК и РНК)
•Синтез ГАГ
08.03.2012 |
168 |
Утилизация пентоз пищи
•В ПЦ вовлекается избыток пищевых пентоз, которые окисляются по пути гликолиза и пр-во АТФ
•Пентозы (пищи)  ПЦ
ПЦ  Гликолиз
Гликолиз
08.03.2012 |
169 |
56

КафедрабиохимииГомГМУ,2012 |
08.03.2012 |
СО2 используется в реакциях:
•биосинтеза ЖК, ГНГ и др.
•регуляции КОС (создание щелочного резерва
крови):
Н2О + СО2  Н2СО3
Н2СО3 Н+ + НСО3-
Н+ + НСО3-
Н+ регулирует [Na+], [K+], [Ca++].
НСО3- регулирует [Cl-]
ПЦ участвует в электрогенезе в нейронах (гиперполяризация -торможение).
08.03.2012 |
170 |
Регуляция ПЦ
•Г6ф-ДГ имеет высокую Км для Г6-ф, поэтому активность ПЦ зависит от [Г6-ф].
Чем она  , тем активнее ПЦ.
, тем активнее ПЦ.
При  [АТФ]
[АТФ]  блок гликолиза
блок гликолиза  Г6-ф активация ПЦ
Г6-ф активация ПЦ
•Активирует  [АТФ], инсулин
[АТФ], инсулин
08.03.2012 |
171 |
Биосинтез ГАГ
•Синтез ГАГ протекает во всех тканях, в том числе и в хрящевой.
•ГАГ состоят из 2 углеводных остатков (димеров):
-Уроновая (идуроновая) кислота,
-N-ацетилглюкозамин (N-ацетилгалактозамин)
08.03.2012 |
172 |
57

КафедрабиохимииГомГМУ,2012 |
08.03.2012 |
Г6-ф
|
Гл-1-ф |
|
|
|
|
|
|
|
|
|
|
Фр-6-ф |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
ГЛН |
УДФ-галактоза |
|
УДФ-глюкоза |
|
|
|
|
|
ГЛУ |
|||||
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
2NAD+ |
Фруктозамин 6-ф |
|
Глюкозамин 6-ф |
||||
|
|
|
|
|
|
2NADH |
|
|
|
|
|||
УДФ-идуроновая |
|
УДФ-глюкуроновая |
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
Фруктозамин-1-ф |
|
N-Ац маннозамин |
||
|
|
|
|
|
|
|
|
|
|
|
|
||
УДФ-ксилоза |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
N-Ац фруктозамин-1-ф |
|
CMP N-Ац |
||||||||
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
УДФ-N-Ац галактозамин |
|
||||||
|
|
|
|
|
|
|
нейраминовая |
||||||
|
|
|
|
|
|
|
УДФ-N- глюкозамин |
|
к-та |
||||
|
|
|
|
|
|
|
|
|
|||||
|
ПУЛ ( pool) |
08.03.2012 |
173 |
ФАФС-фосфоаденозинфосфосульфат
Глюконеогенез – ГНГ
образование глюкозы из неуглеводных субстратов
(глицерина, АК, лактата, ПВК и др.)
•ГНГ снабжает глюкозой прежде всего, мозг и эритроциты.
•ГНГ протекает в в цитоплазме высокоэнергизированных тканей, с большой Мх активностью
•ГНГ это синтетический процесс, требующий большое количество энергии: для синтеза 1 молекулы глюкозы нужно 6 молекул АТФ
08.03.2012 |
174 |
08.03.2012 |
175 |
|
|
58

КафедрабиохимииГомГМУ,2012 |
08.03.2012 |
ГНГ – альтернатива гликолизу
общие обратимые реакции гликолиза и ГНГ:
ГК ФФК
Гл Гл-6ф |
фр-6ф |
ф-1,6 |
4 Г6ф-аза |
3 Ф1.6ф-аза |
|
\ |
|
|
NAD+ NADH |
ПВК киназа |
|
3ФГА 3ФГК
3ФГК  ФЕП
ФЕП  ПВК
ПВК лактат
лактат
NAD+ |
NADH |
АТФ |
1 ПВК |
|
ФЕПКК 2 |
АДФ |
|
|
карбоксилаза |
||
|
|
||
|
|
ОА |
|
08.03.2012 |
|
|
176 |
ПВКкарбоксилаза |
ФЕП-КК |
||
ГНГ: ПВК |
ЩУК |
|
ФЕП |
|
|
|
ДАФ |
2ФГК 3ФГК |
1,3 ДФГК |
3ФГА |
|
Ф1,6-аза |
|
|
Г6Ф-аза |
ф1,6 диф  фр-6ф
фр-6ф  Гл-6ф
Гл-6ф  Гл
Гл
08.03.2012 |
177 |
• Все реакции гликолиза, кроме
гексокиназной, фосфофруктокиназной и пируваткиназной обратимы, поэтому в ГНГ они идут в обратном направлении, с теми же ферментами, что и в гликолизе.
•Эти три необратимые реакции гликолиза, в ГНГ «обращаются» другими специфическими ферментами, отличными от тех, которые катализируют их в гликолизе.
08.03.2012 |
178 |
59
КафедрабиохимииГомГМУ,2012 |
08.03.2012 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
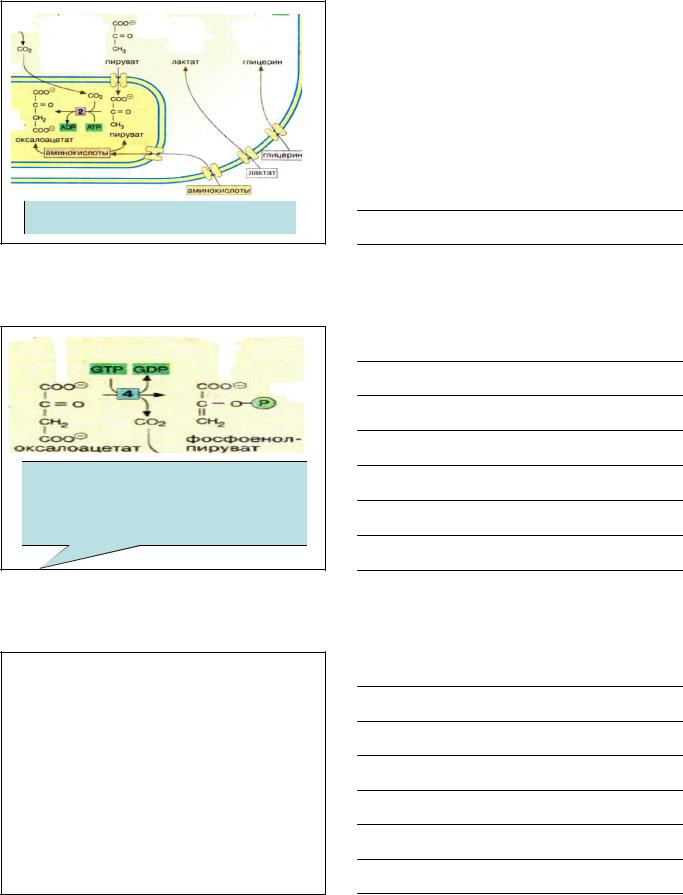
2 -я реакцияПВК--- ЩУК локализуетсяв митохондриях. |
|
Пируваткарбоксилазааллостерический,митохондриальный |
|
|
08.03.2012 |
фермент,активируется ацетил-КоА |
179 |
|
||
Мембрана митохондрий непроницаема для образовавшейся ЩУК, поэтому она восстанавливается в малат, для которого мембрана проницаема. Это связано с тем, что в митохондрии отношение NADH2/NAD относительно велико, поэтому ЩУК легко переходит в малат.
В цитоплазме отношение NADH2/NAD ↓, поэтому малат легко окисляется снова в ЩУК.
08.03.2012 |
180 |
Регуляция ГНГ
и гликолиза реципрокная - теми же факторами, но с обратным знаком.
•факторы, активирующие гликолиз (АМФ, АДФ, глюкоза), ингибируют ГНГ и наоборот
•Факторы ингибирующие гликолиз (АТФ, ЖК, цитрат) активируют ГНГ.
08.03.2012 |
181 |
60