

Торможение в ЦНС. Принципы координационной деятельности ЦНС
..docМинистерство здравоохранения Республики Беларусь
УО «Гомельский государственный медицинский университет»
Кафедра нормальной физиологии
Обсуждено на заседании кафедры
Протокол №__________200__года
ЛЕКЦИЯ №25.
по нормальной физиологии для студентов 2 курса
Тема: Торможение в ЦНС. Принципы координационной деятельности ЦНС.
Время 90 минут
Учебные и воспитательные цели:
Изучить природу, функциональное значение, структурно-функциональную организацию торможения в центральной нервной системе.
ЛИТЕРАТУРА
-
Физиология человека. Под ред. В.М.Покровского, Г.Ф.Коротько.М., Медицина. – 2000.-Т..1 - С 115 – 126.
Мультимедийная презентация 34 слайда.
РАСЧЕТ УЧЕБНОГО ВРЕМЕНИ
|
№ п/п |
Перечень учебных вопросов |
Количество выделяемого времени в минутах |
|
1. |
Введение |
5 |
|
2 |
Торможение в ЦНС. Природа торможения. Виды торможения. |
25 |
|
3 |
Первичное постсинаптическое торможение |
25 |
|
4 |
Пресинаптическое торможение |
10 |
|
5 |
Вторичное торможение |
10 |
|
6 |
Механизмы координации рефлекторной деятельности |
5 |
|
7 |
Компенсация функций и пластичность нервных центров |
5 |
|
8
|
Заключение |
5 |
Всего 90 мин
Торможение в ЦНС. Природа торможения. Виды торможения. Взаимосвязь возбуждения и торможения в ЦНС.
Торможение в ЦНС - особый нервный процесс, вызываемый возбуждением и проявляющийся в подавлении другого возбуждения.
Классификация и механизмы центрального торможения:
1. Первичное постсинаптическое торможение - торможение , не- связанное с первоначальным процессом возбуждения и развивающееся в результате активации специальных тормозных структур. Тормозные синапсы образуют в своих окончаниях тормозной медиатор (ГАМК, глицин, в отдельных синапсах ЦНС роль тормозного медиатора может играть ацетилхолин). На постсинаптической мембране развивается тормозной постсинаптический потенциал (ТПСП), снижающий возбудимость мембраны постсинаптического нейрона. Тормозными нейронами могут служить только вставочные нейроны, афферентные нейроны всегда являются возбуждающими. В зависимости от вида тормозных нейронов и структурной организации нейронной сети постсинаптическое торможение подразделяется на:
Реципрокное торможение. Оно лежит в основе функционирования мышц-антагонистов и обеспечивает расслабление мышцы в момент сокращения мышцы-антагониста. Афферентное волокно, проводящее возбуждение от проприорецепторов мышц (например, сгибателей), в спинном мозге делится на две ветви: одна из них образует синапс на мотонейроне, иннервирующем мышцу-сгибатель, а другая - на вставочном, тормозном, образующем тормозной синапс на мотонейроне, иннервирующем мышцу-разгибатель. В результате возбуждение, приходящее по афферентному волокну, вызывает возбуждение мотонейрона, иннервирующего сгибатель и торможение мотонейрона мышцы-разгибателя.
Возвратное торможение. Оно реализуется через тормозные клетки Реншоу, открытые в спинном мозге. Аксоны мотонейронов передних рогов отдают коллатераль на тормозной нейрон Реншоу, аксоны которого возвращаются на тот же мотонейрон, образуя на нем тормозные синапсы. Таким образом формируется контур с отрицательной обратной связью, позволяющий стабилизировать частоту разрядов мотонейрона.
Центральное (Сеченовское) торможение. Оно осуществляется тормозными вставочными нейронами, через которые реализуется влияние на мотонейрон спинного мозга возбуждения, возникающего в зрительных буграх под влиянием их раздражения. На мотонейроне спинного мозга суммируются ВПСП, возникающие в болевых рецепторах конечности и ТПСП, возникающие в тормозных нейронах под влиянием возбуждения таламуса и ретикулярной формации. В результате время защитного сгибательного рефлекса возрастает.
Латеральное торможение осуществляется с помощью тормозных вставочных нейронов в параллельных нейронных сетях.
Первичное пресинаптическое торможение развивается в терминальных отделах аксонов (перед пресинаптической структурой) под влиянием специальных аксо-аксональных тормозных синапсов. Медиатор этих синапсов вызывает деполяризацию мембраны терминалей и приводит их в состояние, подобное катодической депрессии Вериго. Мембрана в области такого бокового синапса препятствует проведению потенциалов действия к пресинаптической мембране, активность синапса уменьшается.
Пресинаптическое торможение
Пресинаптическим торможением называют снижение или выключение активности клетки за счет синаптического торможения оканчивающейся на ней возбуждающей терминали. Явление пресинаптического торможения зафиксировали Гассер и Грэхем в 1933 г., наблюдая эффект развития торможения сгибательных рефлексов при раздражении других корешков. Данный вид торможения термином “пресинаптическое торможение” впервые обозначили Фрэнк и Фуортес в 1957 г.

Рис. 1. Электронная микрофотография пресинаптических тормозных синапсов. Обозначения: пре. и пост. – пре- и постсинаптический контакт нейрона.
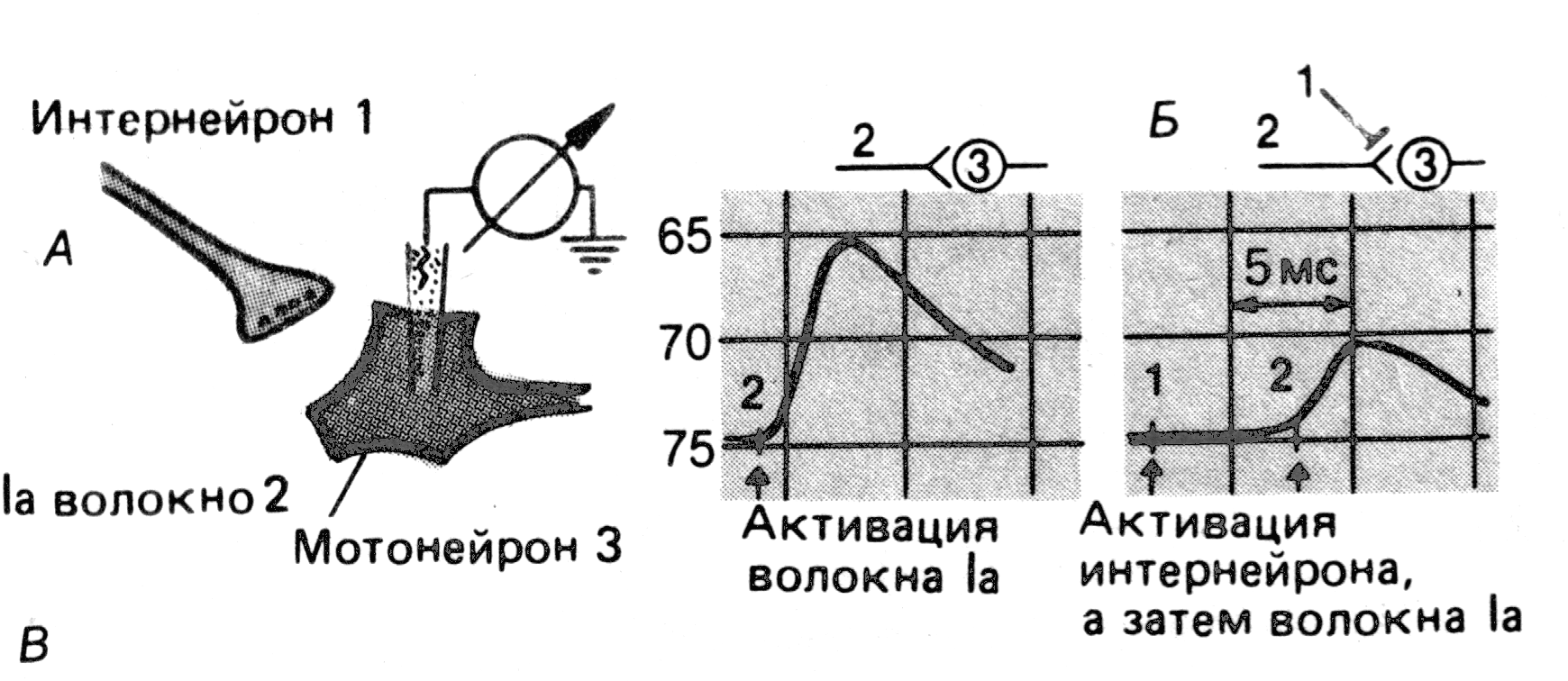
Рис. 2. Пресинаптическое торможение.
А. Схема эксперимента для демонстрации пресинаптического торможения моносинаптических ВПСП в мотонейроне.
Б. ВПСП при стимуляции гомонимных волокон Ia без (слева) и после (справа) предварительной активации пресинаптических тормозных интернейронов.
По “Физиология человека” под ред Р.Шмидта. – 1996. – Т.1. – С.59.
Стимуляция тормозного интернейрона подавляет моносинап-тический ВПСП мотонейрона.
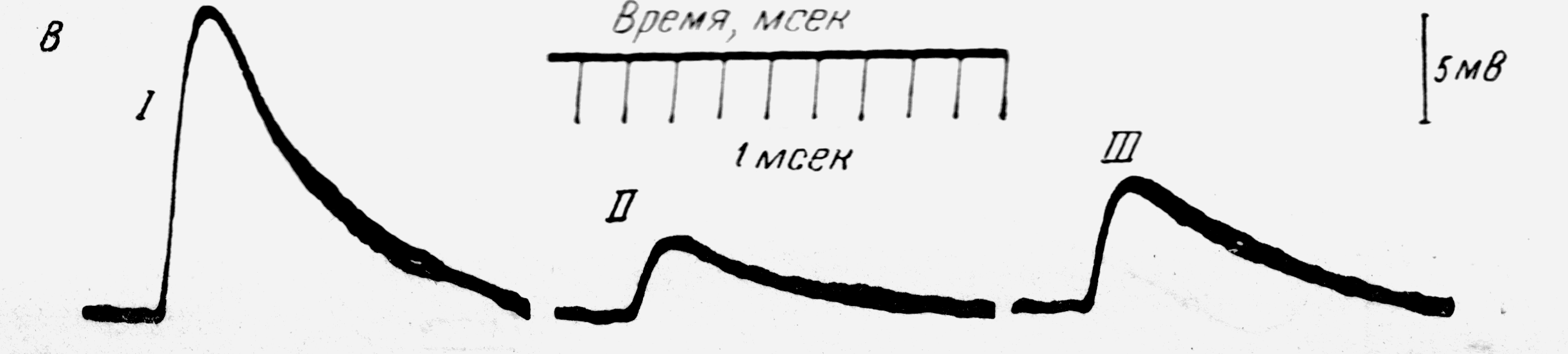
Рис. 3. Подавление моносинаптического ВПСП при синаптическом торможении.
По Дж. Экклс. Физиология синапсов. – М.: Мир. – 1966. – С279.
Развитие этого подавления ВПСП протекает с латентным периодом, равным 5 мс, максимум достигается через 20 мс, а общая длительность превосходит 200 мс.
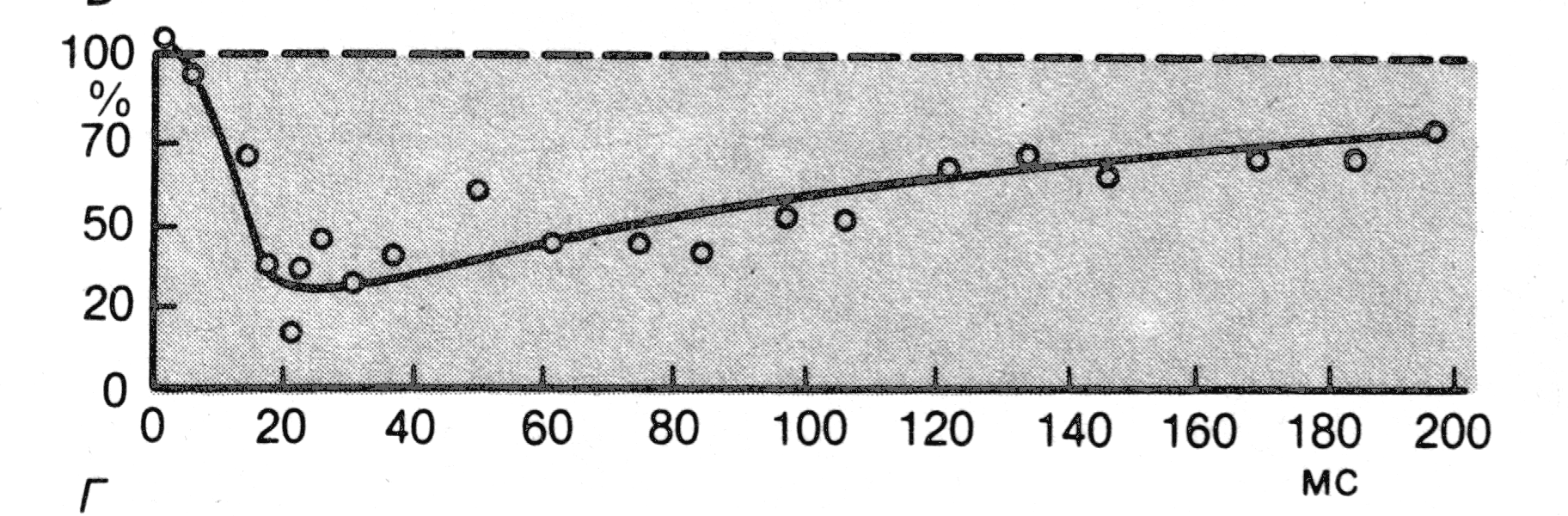
Рис. 4. Временной ход пресинаптического торможения моносинаптических возбуждающих ВПСП.
По “Физиология человека” под ред Р.Шмидта. – 1996. – Т.1. – С.59.
Увеличение частоты предварительных раздражений изменяет характер подавления. В частности, одна серия стимуляции с частотой 200-300 импульсов в секунду вызывает максимальное подавление менее чем на 10%, а две серии – подавление менее чем на 20%. При пресинаптическом торможении подавление моносинаптического ВПСП не связано с какими-либо изменениями их временных параметров.
Тормозные синапсы на окончаниях волокон Ia обеспечивают довольно значительную деполяризацию, называемую деполяризацией первичных афферентов, или же первичную эфферентную деполяризацию (ПАД). В спинном мозгу ПАД обнаруживает длительную фазу (до 25 мс) нарастания до закругленной вершины и характеризуется большей продолжительностью по сравнению с постсинаптическими процессами. Большая продолжительнсоть ПАД объясняется или длительным действием медиатора, или медленным, пассивным снижением деполяризации вследствие большой электрической постоянной времени мембраны. Пассивно снижающаяся компонента ПАД снимается импульсом, распространяющимся по афферентному волокну до его центральных окончаний.
Существует соответствие во всех отношениях между наблюдаемой деполяризацией первичных афферентных волокон и подавлением их синаптического возбуждающего действия.
Пресинаптическая деполяризация афферентов уменьшает величину их пресинаптического спайкового потенциала и таким образом уменьшает вызываемый им ВПСП. По данным Каца (1962), снижение спайкового потенциала на 5 мВ приводит к снижению выброса квантов медиатора и к снижению ВПСП до 50% и менее.
Характер ПАД в различных нейронах отличается по своим характеристикам. В целом временные параметры сравнимы. ПАД волокон кожного нерва отличается большей величиной амплитуды на одиночные раздражения с более коротким латентным периодом (около 2 мс), максимум также достигается ранее, чем в случае ПАД, вызываемых ритмическим раздражением нервных волокон, идущих от мышц. ПАД в клиновидном ядре имеет короткий латентный период (около 2 мс) и быстрый подъем до максимума.
Тормозные синапсы имеют химическую природу, медиатором в них служит ГАМК. Деполяризация первичных афферентов инактивирует возбуждающие натриевые каналы.Шунтирование натриевых каналов снижает амплитуду пресинаптического потенциала действия. В результате синаптическая передача моторного импульса ослабляется или исключается.
Во всех типах возбуждающих синапсов обнаруживается тесная зависимость между деполяризацией пресинаптических волокон и торможением синаптической передачи. Пресинаптическое торможение оказывает влияние на возбуждающие синапсы всех типов, образуемые афферентными волокнами, входящими в спинной мозг, групп Ia, Ib и II, отходящими от мышц, и альфа-волокнами кожного нерва. Это торможение влияет не только на местные спинномозговые рефлексы, но также и на синаптические передачи в восходящих путях как от кожных афферентов, так и на спиноцеребеллярные. Кроме того, пресинаптическое торможение влияет на синаптические передачи задних столбов в ядра нежного и клиновидного пучков. Нисходящие импульсы из коры головного мозга и ствола мозга также оказывают пресинаптическое тормозное влияние на волокна группы I и кожные афферентные волокна в спинном мозгу и клиновидном ядре. Обнаружено пресинаптическое торможение вторичных афферентных волокон отходящих от клиновидного ядра и имеющих переключение в таламусе. Синапсы с пресинаптическим торможением обнаружены в связанном с таламусом ядре мозга – латеральном коленчатом теле. В коре головного мозга не выявлено синаптических структур, которые могли бы осуществлять пресинаптическое торможение. На этих высших уровнях нервной системы доминирует постсинаптическое торможение. Пресинаптическое торможение действует как отрицательная обратная связь, уменьшая приток сенсорной информации в центральную нервную систему. Обычно эта отрицательная обратная связь не имеет точной топографии, но обычно концентрируется в пределах одной сенсорной модальности.
Пресинаптическое торможение служит механизмом регуляции двигательных систем спинного мозга. Его особенностью является возможность специфического воздействия на отдельные синаптические входы без изменений возбудимости всей клетки. Таким образом, избыточная информация устраняется еще до того, как достигает места интеграции клеточного тела нейрона.
2. Вторичное торможение не связано с тормозными структурами, является следствием предшествующего возбуждения. Пессимальное торможение (открыто Н.Е.Введенским в 1886 г.) развивается в полисинаптических рефлекторных дугах при чрезмерной активации центральных нейронов и играет предохранительную роль. Оно выражается в стойкой деполяризации мембраны, приводящей к инактивации натриевых каналов. "Торможение вслед за возбуждением" развивается в нейронах непосредственно после потенциала действия и характерно для клеток с длительной следовой гиперполяризацией. Таким образом, процессы торможения в локальных нейронных сетях уменьшают избыточную активность и участвуют в поддержании оптимальных режимов активности нейронов.
-
Механизмы координации рефлекторной деятельности: реципрокная иннервация, доминанта (А.А.Ухтомский), принципы обратной связи и общего конечного пути, принцип субординации.
Принцип иррадиации возбуждения. Иррадиация - распространение, расширение рефлекторного ответа. Это феномен “растекания” возбуждения по нейронам центральной нервной системы, развивающийся или после действия сверхсильного раздражителя, или на фоне выключения торможения. Распространение возбуждения возможно за счет многочисленных контактов между нейронами, возникающих при ветвлении аксонов и дендритов вставочных нейронов. Иррадиация позволяет увеличивать количество участвующих в рефлекторном ответе групп мышц. Ограничивают иррадиацию тормозные нейроны и синапсы.
На фоне действия стрихнина, блокирующего тормозные синапсы, наступают генерализованные судороги при тактильной стимуляции любого участка тела или при раздражении рецепторов любой сенсорной системы. В коре больших полушарий наблюдается явление иррадиации процесса торможения.
В основе координации рефлекторных актов лежат определенные механизмы, основанные на структурно-функциональной организации ЦНС и обозначаемые как “принципы” формирования рефлекторного ответа.
Принцип реципрокной иннервации. Реципрокная (сопряженная) координация открыта Н.Е.Введенским в 1896 году. Обусловлена реципрокным торможением, т.е. активация одного рефлекса одновременно сопровождается торможением второго, противоположного по своей физиологической сущности.
Принцип общего "конечного пути". Открыт английским физиологом Ч.Шеррингтоном (1906). Один и тот же рефлекс (например, сокращение мышцы) может быть вызван раздражением различных рецепторов, т.к. один и тот же конечный -мотонейрон передних рогов спинного мозга входит в состав многих рефлекторных дуг. Рефлексы, дуги которых имеют общий конечный путь, подразделяются на агонистические и антагонистические. Первые усиливают, вторые тормозят друг друга, как бы конкурируя за конечный результат. В основе подкрепления лежит конвергенция и суммация, в основе конкуренции за конечный путь - сопряженное торможение.
Принцип обратной связи. Любой рефлекторный акт контролируется благодаря обратной связи с центром. Обратная связь состоит во вторичной афферентации, поступающей в ЦНС от рецепторов, которые возбуждаются при изменении функциональной активности рабочего органа. Например, потенциалы действия, обусловленные возбуждением рецепторов мышц, сухожилий и суставных сумок сгибающейся конечности, в процессе осуществления акта сгибания поступают во все структуры ЦНС, начиная от центров спинного мозга. Различают обратную связь положительную (усиливающую рефлекс, который является источником обратной афферентации) и отрицательную, когда рефлекс, ее вызывающий, тормозится. Обратная связь лежит в основе саморегуляции функций организма.
Принцип отдачи. Феномен отдачи состоит в быстрой смене одного рефлекса другим противоположного значения. Например, после сгибания конечности ее разгибание происходит быстрее, особенно если сгибание было сильным. Механизм этого явления состоит в том, что при сильном сокращении мышц возбуждаются рецепторы Гольджи сухожилий, которые через тормозные вставочные нейроны тормозят мотонейроны сгибательных мышц и образуют ветвь, которая возбуждает центр мышц - разгибателей. Благодаря этому механизму можно получить сумму рефлексов - цепные рефлексы (окончание одного рефлекторного ответа инициирует следующий) и ритмические (многократное повторение ритмичных движений).
Принцип доминанты. Конечный поведенческий эффект при координации рефлексов может быть изменен в зависимости от функционального состояния центров (наличия доминантных очагов возбуждения).
Особенности доминантного очага возбуждения:
1. Повышенная возбудимость нейронов.
2. Стойкость процесса возбуждения.
3. Способность к суммации возбуждения.
4. Иннертность. Очаг доминирует, подавляет соседние центры путем сопряженного торможения, возбуждаясь за их счет. Доминанту можно получить химическим воздействием на центры, например, стрихнином. В основе доминантного возбуждения лежит способность возбудительного процесса к иррадиации по нейронным цепям.
Компенсация функций и пластичность нервных центров
Компенсаторная функция нервных центров заключается в том, что даже после значительных разрушений части нервных клеток и глиальных элементов происходит восстановление функций центра. В отдельных случаях центр способен перестроиться таким образом, что может принимать участие в регуляции совершенно иной функции.
Пластичность - это способность нервных центров изменять свое прямое функциональное назначение и расширять свои функциональные возможности. Это качество генетически запрограммировано, но может значительно развиваться под влиянием упражнений. Пластичность ЦНС проявляется при травмах, когда функция поврежденного участка (например, коры больших полушарий головного мозга) обеспечивается за счет сохранившихся или смежных отделов.
Заведующий кафедрой, профессор Э.С. Питкевич
Дата переработки 8 декабря 2006г.