

Лекция 8.
Мембранные потенциалы и их ионная природа. Потенциал покоя. Уравнение Нернста. Уравнение Гольдмана-Ходжкина-Катца.
Механизм генерации потенциала действия. Распространение потенциала действия по миелиновым и безмиелиновым нервным волокнам.
11.7. Равновесный и стационарный мембранные потенциалы. Потенциал покоя
Опыты Л. Гальвани и А. Вольта во второй половине XVIII в. привели к пониманию того, что функционирование живых тканей сопровождается электрическими явлениями. В настоящее время неоспоримым является тот факт, что генерация и распространение электрических потенциалов — это важнейшее физическое явление в живых клетках и тканях.
Биопотенциалом называют разность электрических потенциалов, образующуюся между двумя точками клеток, тканей и органов в процессе их жизнедеятельности. Биопотенциалы отражают функциональное состояние клеток и тканей. Поэтому их регистрация и анализ являются важным приемом при физиологических исследованиях и в диагностике.
Д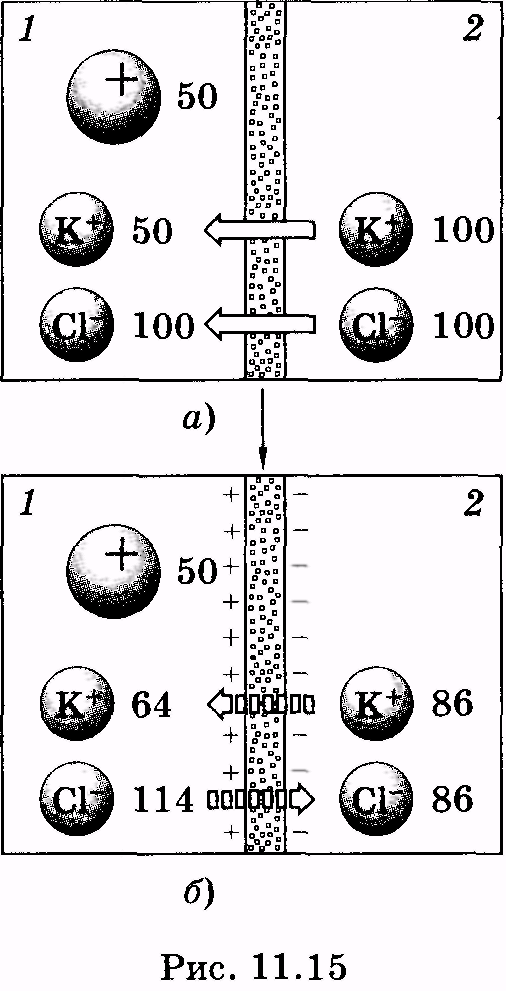 ля
понимания природы мембранных потенциалов
— электрических потенциалов,
образующихся между внутренней и внешней
сторонами мембраны, рассмотрим сначала
модельную систему,представляющую
собой сосуд, разделенный полупроницаемой
мембраной (рис.
11.15). Предположим, в левой
части сосуда содержатся ионы калия,
хлора и какие-либо крупные частицы,
например молекулы белка, несущие
положительный заряд (раствор
1).
В
правой части сосуда находятся
только калий и хлор (раствор 2).
Мембрана
способна легко пропускать неорганические
анионы и катионы, но является
непроницаемой для молекул белка.
Цифрами указаны относительные
концентрации соответствующих ионов,
при которых разность потенциалов
составит около 10 мВ.
ля
понимания природы мембранных потенциалов
— электрических потенциалов,
образующихся между внутренней и внешней
сторонами мембраны, рассмотрим сначала
модельную систему,представляющую
собой сосуд, разделенный полупроницаемой
мембраной (рис.
11.15). Предположим, в левой
части сосуда содержатся ионы калия,
хлора и какие-либо крупные частицы,
например молекулы белка, несущие
положительный заряд (раствор
1).
В
правой части сосуда находятся
только калий и хлор (раствор 2).
Мембрана
способна легко пропускать неорганические
анионы и катионы, но является
непроницаемой для молекул белка.
Цифрами указаны относительные
концентрации соответствующих ионов,
при которых разность потенциалов
составит около 10 мВ.
Общее число частиц в растворах одинаково (сохраняется их электронейтральность), однако концентрация ионов калия во втором растворе больше (см. рис. 11.15, а). Ионы калия устремятся из раствора 2 в раствор 1, а вслед за ними пойдут и ионы хлора (для сохранения электронейтральности растворов). При этом концентрация ионов хлора в первом растворе еще больше возрастет. Этот процесс будет продолжаться до установления равновесного состояния (так называемого равновесия Доннана). Между двумя сторонами мембраны образуется разность потенциалов, которая уравновешивает концентрационный градиент ионов, способных к диффузии (на рис. 11.15, б потоки соответствующих ионов указаны штриховыми стрелками). Мембранная разность потенциалов рассчитывается по формуле Нернста:
![]() (11.35)
(11.35)
Здесь с1 и с2 — молярные концентрации ионов по обе стороны мембраны, R — универсальная газовая постоянная, Т — термодинамическая температура, при которой происходит диффузия, F — постоянная Фарадея, Z — заряд иона. Эту разность потенциалов называют равновесным мембранным потенциалом. Мембранная теория происхождения биопотенциалов была выдвинута в 1902 г. Б. Бернштейном. Действительно, в живой клетке концентрация ионов калия значительно больше, чем в межклеточной жидкости, и крупные органические молекулы практически не проникают через мембрану. Важным доводом в пользу представлений Бернштейна послужил тот факт, что рассчитанная по формуле Нернста разность потенциалов между наружной и внутренней сторонами мембраны мышечного волокна оказалась близкой к измеренной в опытах с помощью внутриклеточного микроэлектрода.
Однако «калиевая теория» мембранного потенциала оказалась несовершенной, не способной объяснить наблюдаемые впоследствии факты отклонения истинных значений потенциалов на мембранах живых клеток от теоретически рассчитанных. Оказалось, что равновесный мембранный потенциал характерен лишь для мертвых клеток, либо клеток с ослабленным метаболизмом.
В настоящее время общепризнанной теорией, объясняющей возникновение и поддержание потенциала на клеточной мембране в состоянии физиологического покоя, является теория А. Ходжкина. Она была развита и экспериментально обоснована им в 50-х гг.XXв. Сущность ее заключается в том, что потенциал, существующий на мембранах невозбужденных клеток (потенциал покоя), обусловлен полупроницаемыми свойствами клеточной мембраны и неравномерным распределением ионов между клеткой и окружающей средой. Это распределение поддерживается механизмами активного переноса, локализованными в самой мембране.
При получении выражения для потенциала покоя важно учитывать знаки ионов, проникающих через мембрану. Это можно сделать, в частности, обозначая знаком плюс плотности потоков положительных ионов и знаком минус — отрицательных. Основной вклад в создание и поддержание потенциала покоя вносятионы натрия, калия и хлора. Суммарная плотность потока этих ионов с учетом их знаков равна
![]() (11.36)
(11.36)
Для живой клетки характерно не равновесное распределение веществ, но существование потоков ионов в обе стороны через мембрану. Такое состояние, при котором число различных ионов, проходящих в единицу времени через мембрану внутрь клетки, равно числу выходящих из клетки ионов, называют стационарным. Ясно, что в стационарном состоянии суммарная плотность потока ионов через мембрану равна нулю: J = 0.
Для плотности потоков положительных ионов натрия и калия и отрицательных ионов хлора запишем общее выражение на основании (11.33) и (11.34)
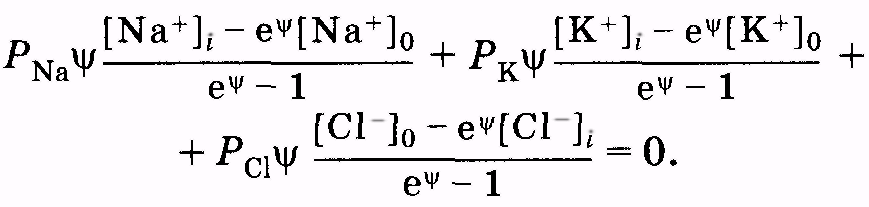 (11.37)
(11.37)
Здесь
квадратными скобками [ ]i,
и [ ]0
обозначены концентрации ионов
соответственно внутри и вне клетки.
Сократив (11.37) на
![]() ,
раскрыв выражения и перегруппировав
их, получаем
,
раскрыв выражения и перегруппировав
их, получаем
PNa[Na+]i + РК[К+]i + РСl[Сl-]0 = e {PNa[Na+]0 + РК[К+]0 + РС1[Сl-] i},
или
![]()
Логарифмируя это выражение, находим
![]() (11.38)
(11.38)
Если от безразмерного потенциала вернуться к электрическому потенциалу [см. (11.29)], то из (11.38) получаем
![]() (11.39)
(11.39)
— уравнение Гольд мана — Ходжкина — Катца.
Различные концентрации ионов внутри и вне клетки созданы ионными насосами — системами активного транспорта. Можносказать, что потенциал покоя обязан активному переносу.
Для большей наглядности рассмотрим схематично ионные потоки, существующие в состоянии покоя на мембране аксона кальмара (рис. 11.16). На рисунке жирными стрелками указаны потоки ионов, осуществляемые за счет активного транспорта. Пунктиром обозначены стрелки, соответствующие диффузионным потокам засчет градиентов концентрации соответствующих ионов. Обозначены также концентрации ионов натрия, калия и хлора в цитоплазме и окружающей среде.
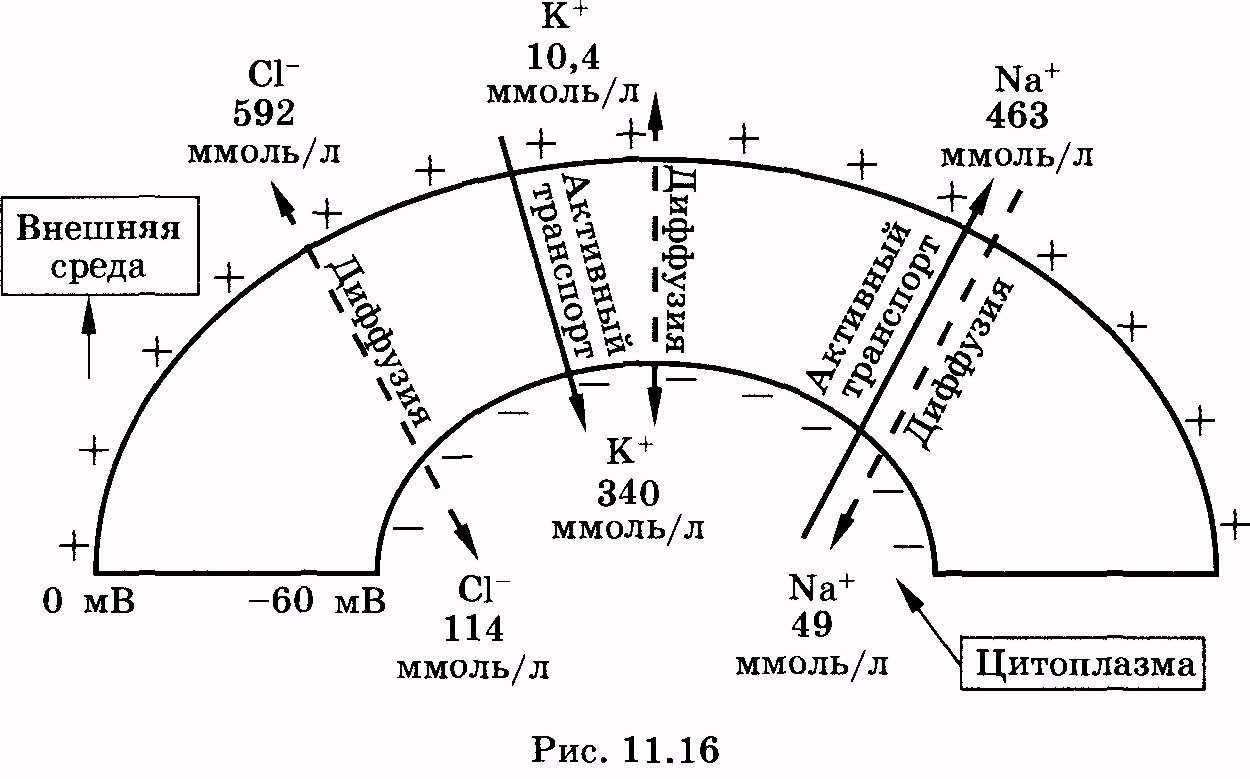
Известно, что проницаемость мембраны для ионов калия самая высокая. В состоянии покоя соотношение коэффициентов проницаемости для разных ионов равно:
![]()
Вследствие этого диффузия калия и хлора идет в обе стороны. Натрий идет через мембрану за счет простой диффузии лишь в одну сторону — снаружи вовнутрь. Однако Ма+-К+-АТФаза интенсивно выводит ионы натрия из клетки, а калия — в клетку. Потенциал покоя, рассчитанный по формуле Гольдмана—Ходжкина— Катца, составляет 60 мВ со знаком минус со стороны внутриклеточного пространства.
На основании большого экспериментального материала было установлено, что величина потенциала покоя может существенно различаться для разных клеток. В таблице 19 приведены данные для различных тканей.
Таблица 19. Потенциал покоя клеточных мембран для различных тканей
|
Ткань |
Потенциал покоя, мВ |
|
Аксон кальмара Нерв лягушки Поперечно-полосатое мышечное волокно лягушки Сердечное мышечное волокно лягушки Сердечное волокно собаки Клетки водорослей |
60 70 88 70 90 100-120 |