11.Размножение - универсальное свойство живого, обеспечивающее материальную непрерывность в ряду поколений. Эволюция размножения, формы размножения.
Размножение – это способность организмов производить себе подобных особей того же вида. Существует два типа размножения: половое и бесполое. Древнейшим способом размножения на Земле было бесполое размножение. Бесполое размножение. Бесполое размножение происходит без образования специальных клеток, в нём участвует один организм, одна особь, при этом размножении образуются идентичные потомки. Единственным источником генетической изменчивости являются случайные мутации. Цитологической основой бесполого размножения является митоз. Молекулярной основой бесполого размножения является репликация ДНК. Бесполое размножение у различных живых организмов может происходить по-разному. Формы бесполого размножения: 1. Почкование – это форма бесполого размножения при которой новая особь образуется в виде выростов (почки) на теле родительской особи, а затем отделяется от неё и превращается в самостоятельную особь (гидра, дрожжи). 2. Фрагментация – это разделение особи на две или более частей, каждая из которых растёт и образуется отдельная особь (высшие растения, губка, дождевой червь). 3. Образование спор. Спора – это одноклеточная репродуктивная единица, состоящая из ядра и небольшого количества цитоплазмы под плотной оболочкой. Из споры образуется новая особь (низшие растения). 4. Деление. Бинарное деление клетки на две части. Ядро родительской особи один или несколько раз делится митозом, при этом образуется два или несколько дочерних ядер. Каждое из них окружается цитоплазмой и развивается в самостоятельный организм. 5. Шизогония – это множественное деление клетки. Сначала в клетке многократно делится ядро, затем вокруг каждого ядра обособляется участок цитоплазмы, который окружается плазматической мембраной. Затем происходит распад на отдельные клетки (малярийный плазмодий). 6. Вегетативное размножение. Осуществляется формирование дочернего организма из группы клеток материнского организма. У растений это размножение происходит за счёт вегетативных органов: корневищ, луковиц, клубней, усов. В результате бесполого размножения образуются генетически идентичные особи. Скорость размножения очень высокая и в постоянных условиях организма быстро захватывают экологическую нишу. Половое размножение. Появилось половое размножение более 3 млрд. лет назад. Сущность полового размножения в перекомбинации генетического материала родительских особей. В результате дочерние особи становятся более разнообразными, и естественный отбор выбирает из них наиболее приспособленные. При половом размножении потомство получается в результате слияния гаплоидных клеток – гамет. При оплодотворении образуется зигота. Из которой развивается новый организм. Оплодотворение – это процесс слияния сперматозоида с яйцеклеткой с последующим слиянием их ядер и образованием диплоидной зиготы. Биологическое значение этого процесса состоит в том, что при слиянии мужских и женских гамет образуется новый организм, несущий признак обоих родительских организмов. Гаметы гаплоидны, они содержат половинный набор хромосом и образуются в результате мейоза. Одной из модификаций полового размножения является партеногенез. Партеногенез – это процесс, при котором женская гамета развивается в новую особь без оплодотворения (встречается у животных (пчёлы) и растений). Преимущество в том, что увеличивается скорость размножения.
Эволюция размножения стала важной движущей силой общей эволюции царства растений и привела к возникновению новых специализированных органов. Бесполое размножение — наиболее древний и, несомненно, изначальный способ размножения. Оно представлено у всех групп растений, включая покрытосеменные
Наиболее примитивный способ бесполого размножения — вегетативное. Его наблюдают на самых ранних этапах развития жизни — у доклеточных. У многих предъядерных (например, у бактерий) это и сейчас единственный способ размножения. Каждая крупная естественная группа растений нередко обладает свойственными только ей формами вегетативного размножения.
Так, у сине-зеленых водорослей это гормогонии, у лишайников — изидии и соредии, у грибов — хламидоспоры и почкование, у некоторых моховидных— выводковые тельца. Наиболее разнообразны формы вегетативного размножения у высших растений, особенно у покрытосеменных. Велико разнообразие спор собственно бесполого размножения. В процессе эволюции они утратили способность воспроизводить новую особь, сходную с материнской (спорофит), а дают начало половому поколению (гаметофиту). Половое размножение возникло на очень ранних этапах эволюции, но время его появления точно не установлено.
У некоторых современных примитивных групп растений половой процесс неизвестен. Не вызывает сомнений, что такие природные группы, как сине-зеленые водоросли и большинство бактерий, никогда не имели полового размножения. Другие, например некоторые грибы, возможно, утратили его. Половой процесс утрачен и у некоторых высокоорганизованных растений, например у покрытосеменных (партеногенез). В настоящее время наука еще не располагает достаточными данными для воспроизведения полной картины эволюции полового процесса у растений.
Твердо установлено, что у растений, стоящих на более низкой ступени эволюции, половой процесс протекает в более примитивной форме, чем у растений, стоящих на более высокой ступени эволюции. Один из показателей уровня эволюции — специализация гамет. Наиболее примитивен изогамный половой процесс. Более высокий уровень — гетерогамия.
Наиболее специализированным половым процессом считают оогамию. Мужская гамета теряет подвижность, приспособившись к наземным условиям жизни, и доставляется к яйцеклетке пыльцевой трубкой. Таким образом, у семенных растений половой процесс не связан с наличием водной среды, это — приспособление к сухопутному образу жизни. Половое размножение развивалось не изолированно от бесполого, а во взаимной связи с ним. Каждая природная группа растений в жизненном цикле имеет свою закономерность смены бесполого и полового размножения. У многих низших растений смена ядерных фаз не выражена, спорофит почти полностью выпадает из жизненного цикла (диплоидна лишь зигота).
В жизненном цикле некоторых низших растений и всех высших существует чередование спорофита и гаметофита. Степень их морфологической разнородности и преобладания в жизненном цикле служит важным показателем эволюционного уровня изучаемой группы растений. На основании работ В. Гофмейстера (1849, 1851) установлена важная общая закономерность: в процессе эволюции спорофит приобретает всё большее развитие и самостоятельность; гаметофит, напротив, все более редуцируется, полностью теряет свою самостоятельность и всецело зависит от спорофита, что и наблюдают у наиболее высокоорганизованных семенных растений — голосеменных и покрытосеменных. В. Гофмейстер, изучая жизненные циклы различных природных групп растений, показал, что между высшими споровыми и покрытосеменными нет такого большого разрыва, как предполагали ранее.
12,13. Овогенез. Характеристика основных периодов. Строение женских половых гонад.
Сперматогенез. Характеристика основных периодов. Строение мужских половых гонад.
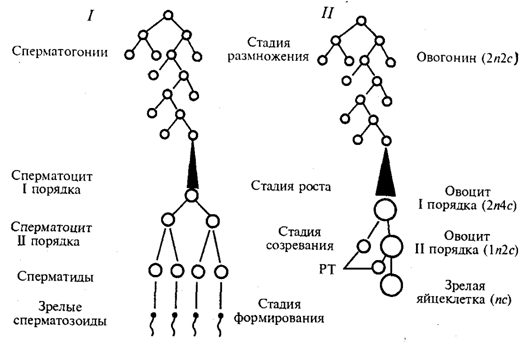
Гаметогенез — процесс образования яйцеклеток (овогенез) и сперматозоидов (сперматогенез) —подразделяется наряд стадий (рис. 5.4).
В стадии размножения диплоидные клетки, из которых образуются гаметы, называют сперматогониями и овогониями. Эти клетки осуществляют серию последовательных митотических делений, в результате чего их количество существенно возрастает. Сперматогонии размножаются на протяжении всего периода половой зрелости мужской особи. Размножение овогоний приурочено главным образом к периоду эмбриогенеза. У человека в женском организме этот процесс наиболее интенсивно протекает в яичниках между 2-м и 5-м месяцами внутриутробного развития. К 7-му месяцу большая часть овоцитов входит в профазу I мейоза.
Так как способом размножения клеток-предшественниц женских и мужских гамет является митоз, то овогоний и сперматогонии, как и все соматические клетки, характеризуются диплоидностью. В ходе митотического цикла их хромосомы имеют либо однонитчатую (после митоза и до завершения синтетического периода интерфазы), либо двунитчатую (постсинтетический период, профаза и метафаза митоза) структуру в зависимости от количества биспиралей ДНК. Если в одинарном, гаплоидном наборе число хромосом обозначить как п, а количество ДНК — как с, то генетическая формула клеток в стадии размножения соответствует 2п2с до S-периода и 2n4c после него.
Рис. 5.4. Схема гаметогенеза:
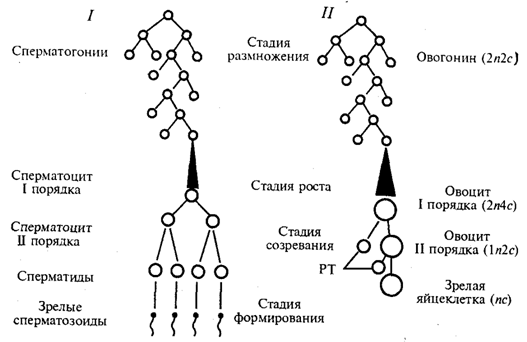
1 — сперматогенез, 2 — овогенез, n — количество хромосомных наборов,с — количество ДНК, РТ — редукционные тельца
На стадии роста происходит увеличение клеточных размеров и превращение мужских и женских половых клеток в сперматоциты и овоциты I порядка, причем последние достигают больших размеров, чем первые. Одна часть накапливаемых веществ представляет собой питательный материал (желток в овоцитах), другая — связана с последующими делениями. Важным событием этого периода является репликация ДНК при сохранении неизменным числа хромосом. Последние приобретают двунитчатую структуру, а генетическая формула сперматоцитов и овоцитов I порядка приобретает вид 2n4с.
Основными событиями стадии созревания являются два последовательных деления: редукционное и эквационное, которые вместе составляют мейоз (см. разд. 5.3.2). После первого деления образуются сперматоциты и овоциты II порядка (формула n2с), а после второго — сперматиды и зрелая яйцеклетка (пс).
В результате делений на стадии созревания каждый сперматоцит I порядка дает четыре сперматиды, тогда как каждый овоцит I порядка — одну полноценную яйцеклетку и редукционные тельца, которые в размножении не участвуют. Благодаря этому в женской гамете концентрируется максимальное количество питательного материала — желтка.
Процесс сперматогенеза завершается стадией формирования, или спермиогенеза. Ядра сперматид уплотняются вследствие сверхспирализации хромосом, которые становятся функционально инертными. Пластинчатый комплекс перемещается к одному из полюсов ядра, образуя акросомный аппарат, играющий большую роль в оплодотворении. Центриоли занимают место у противоположного полюса ядра, причем от одной из них отрастает жгутик, у основания которого в виде спирального чехлика концентрируются митохондрии. На этой стадии почти вся цитоплазма сперматиды отторгается, так что головка зрелого сперматозоида практически ее лишена.
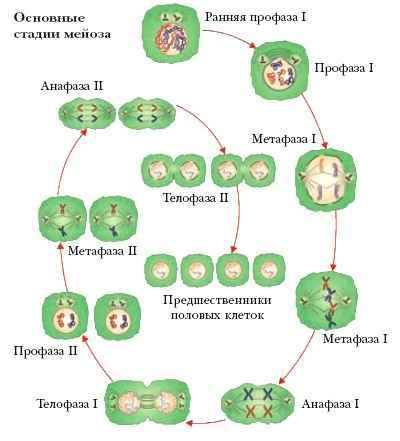
15.Мейоз
МЕЙОЗ (деления созревания, период созревания), этап в образовании половых клеток; состоит из двух последовательных делений исходной диплоидной клетки (содержат два набора хромосом – 2n) и формирования четырёх гаплоидных половых клеток, или гамет (содержат по одному набору хромосом – n). Уменьшение (редукция) числа хромосом (2nn) происходит за счёт того, что на два деления приходится лишь одно удвоение (репликация) хромосомного материала. При оплодотворении гаплоидные гаметы – яйцеклетка и сперматозоид – сливаются и диплоидное число хромосом, характерное для каждого вида, восстанавливается (n + n2n). В главных чертах мейоз протекает сходно у разных групп организмов и у особей женского и мужского пола. Два следующих друг за другом деления первичной половой клетки обозначаются как мейоз I и мейоз II. Подобно делению соматических клеток – митозу, и мейоз I, и мейоз II состоят из четырёх основных стадий – профазы, метафазы, анафазы и телофазы. Вступающая в мейоз клетка диплоидна, а каждая хромосома содержит удвоенное количество ДНК. В первом мейотическом делении особенно сложна и длительна профаза I (у человека она занимает 22,5 сут). На этой стадии гомологичные хромосомы соединяются (конъюгируют) в пары – биваленты. В каждой хромосоме бивалента различимы в микроскопе две продольные половины – хроматиды, т. е. бивалент представляет собой четвёрку (тетраду) хроматид. В профазе I происходит генетически значимое событие – обмен гомологичными (содержащими одни и те же гены) участками несестринских хроматид, или кроссинговер. В анафазе I биваленты разъединяются и гомологичные хромосомы расходятся к противоположным полюсам клетки, причем, в отличие от анафазы митоза, каждая хромосома сохраняет две хроматиды. В результате число хромосом уменьшилось вдвое, но удвоенным остаётся и количество ДНК, представленное двумя хроматидами. Важная особенность расхождения хромосом заключается в том, что любая, отцовская или материнская, хромосома из гомологичной пары может отойти к любому из полюсов независимо от того, как расходятся хромосомы других пар. Это означает, что число возможных сочетаний хромосом в дочерних клетках обычно очень велико: 2n, где n – число хромосомных пар (у человека – 223). Так происходит ещё одно перемешивание родительского генетического материала – рекомбинация хромосом. После мейоза I обычно сразу или после короткой интерфазы, во время которой удвоение хромосом не происходит, следует мейоз II. Это деление аналогично митозу с той разницей, что делятся гаплоидные клетки. В анафа-зе II сестринские хроматиды разделяются и, став хромосомами, расходятся к полюсам. Число хромосом и количество ДНК приходят в соответствие, и мейоз II завершается образованием четырёх гаплоидных гамет, каждая из которых несёт уникальный генетический материал. У самок, однако, лишь одна из четырёх гамет – яйцеклетка, способная к оплодотворению. Мейоз – один из ключевых биологических процессов. Его значение состоит в поддержании в поколениях постоянства хромосомных наборов (кариотипов), т. е. в обеспечении наследственности, и в создании новых сочетаний отцовских и материнских генов, т. е. в обеспечении генотипической изменчивости.