

Хондроитинсульфаты (хс). Это полимеры, структурной единицей которых является димер, состоящий из глюкуроновой кислоты и n-ацетилгалактозамина (сульфатирован по 4 или 6 положению).
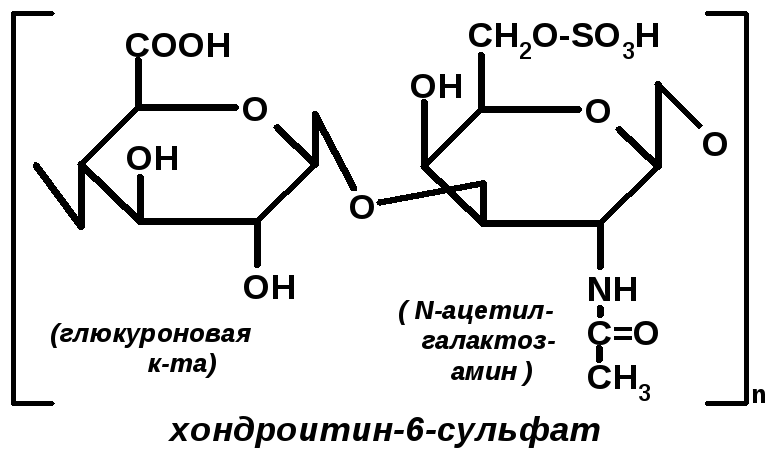
ХС-ты входят в состав опорной, костной ткани, хрящей, сухожилий. Наличие сульфатной группировки и ОН-группы глюкуроновой кислоты придаёт этим соединениям дополнительные отрицательные заряды в растворе, и эти полимеры являются полианионами. Поэтому они легко притягивают Са2+ и таким образом в процессах минерализации играют определённую роль в связывании Са. В межклеточном матриксе протеогликаны организованы в своеобразные комплексы («ерш из ершиков»). Такой комплекс содержит до 160 протеогликанных единиц.
Гепарин. Полимер, структурной единицей является димер, состоящий из глюкуроновой кислоты, связанной с N-ацетилглюкозамином, сульфатированым 4 или по 6 положению серной кислотой.

Гепарин не является структурным компонентом межклеточного матрикса. Он вырабатывается тучными клетками соединительной ткани и после их цитолиза выделяется в межклеточное пространство и кровеносное русло. В крови может соединяться с неспецифическими белками. Гепарин – естественный природный коагулянт, препятствует свёртыванию крови.
ПГ выполняют следующую роль:
1. роль «рессор» – смягчают нагрузки на суставные поверхности (в хрящах, суставных поверхностях);
2. ограничивают диффузию и проницаемость межклеточного вещества для различных молекул и крупных частиц. Гиалуроновая кислота подвергается гидролизу под действием фермента – гиалуронидазы. В норме, в здоровой ткани, фермент и субстрат находятся в равновесии. при патологических процессах количество фермента может увеличится, что приводит к гидролизу гиалуроновой кислоты и нарушению проницаемости. При этом разрываются -1,4-гликозидные связи;
3. являются поливалентными анионами. Способны связывать большие количества ионов Na+ и др. катионов и участвуют в регуляции водно-солевого обмена;
4. Гиалуроновая кислота может присутствовать в свободном виде, например в хрусталике глаза, суставной жидкости. Она имеет консистенцию геля и выполняет различные функции.
Фосфопротеины
Белки, где в качестве простетической группы – фосфорная кислота. Присоединение фосфорной кислоты к полипептидной цепи идет с образованием сложноэфирной связи с АК СЕР или ТРЕ.
![]()
![]()
Типичными фосфопротеинами являются: казеин молока (1% фосфорной кислоты), желток куриного яйца (вителлин), икра рыб (ихтулин). Большое количество ФП содержится в нервных клетках. Для ФП характерен широкий диапазон функций в метаболизме. За счет фосфорилирования и дефосфорилирования ферментов происходит регуляция их активности.
Ферменты
Ферменты – это биологические катализаторы белковой природы. В одной клетке до 10 тыс. молекул фермента, которые катализируют 2000 ферментативных реакций. 1800 тыс. ферментов выделены, но их строение не расшифровано. Старое название ферментов – энзимы, а наука, их изучающая – энзимология.
По своей химической природе ферменты – это белки, они имеют несколько уровней структурной организации и обладают всеми другими свойствами белков. Очень большое количество ферментов не имеют 4-ую структуру, т.е. являются олигомерами.
Ферменты могут быть простыми и сложными. Простые состоят только из полипептидной цепи, а сложные имеют пептидную (апофермент) и небелковую части (кофермент). Далее идёт рис. [апофермент+кофермент=холофермент – обладает полноценной функциональной активностью]. По отдельности ни апофермент, ни кофермент, не выполняют функции так, как холофермент.
Строение ферментов. В пространственной структуре фермента условно выделяют ряд участков, которые выполняют соответствующие им функции. Активный центр (АЦ) – участок в молекуле фермента, где происходит связывание и химическое превращение субстрата (S). Субстрат – вещество, подвергающееся химическому превращению (например, для фермента лактатдегидрогеназы (ЛДГ) субстратом будет молочная кислота). В активном центре выделяется контактный участок и каталитический участок. Контактный участок – это место активного центра, в котором происходит связывание фермента с субстратом по принципу комплементарности, т.е. именно контактный участок обеспечивает специфическое сродство субстрата ферменту. Образовавшийся комплекс носит название фермент-субстратный комплекс. Каталитический участок (центр) – это место в активном центре фермента, где происходит химическое превращение субстрата [рис. изображён фермент-субстратный комплекс, а именно контактный участок, каталитический участок, активный центр и субстрат].
Если фермент – сложный белок, то обычно простетическая часть находится тоже в активном центре и участвует в формировании активного центра. Активный центр занимает небольшую часть молекулы фермента, обычно располагается в углублении, и в его образовании участвует небольшое число аминокислотных остатков (до 20). Аминокислотные остатки могут быть удалены друг от друга, но при формировании пространственной структуры фермента они располагаются в области активного центра.
В формировании активного центра могут участвовать остатки, несущие следующие функциональные группы: NH2 (ЛИЗ, АРГ), COOH (ГЛУ, АСП), OH (СЕР, ТРЕ), SH (ЦИС), имидазольное кольцо (ГИС). В качестве единиц, участвующих в формировании активного центра, могут выступать кофакторы - ионы металлов (Cu2+, Fe2+ и т.д.), а также коферменты. В сложном ферменте АК-остатки активного центра создают условия для правильной его конформации и помогают кофакторам в связывании, ориентации, а, следовательно, и в превращении субстрата.
Боковые группы остальных аминокислот не участвуют в образовании активного центра, но обеспечивают правильную пространственную конформацию активного центра и влияют на его реакционную способность.
Ряд ферментов могут содержать аллостерический центр. [рис. фермента с аллостерическим и активным центрами] Эти ферменты относят к аллостерическим ферментам. К аллостерическому центру присоединяются различные вещества, отличные по строению от субстрата. Эти вещества могут изменять конформацию активного центра, т.е. влиять на связывание и превращение субстрата, они называются аллостерическими эффекторами. Все аллостерические эффекторы делятся на положительные – активаторы, и отрицательные – ингибиторы.