

Лекция_Строение_Мембраны
.pdfЛекция
СТРОЕНИЕ БИОМЕМБРАН
Мембранные структуры клеток
Успехи в исследовании мембран во многом достигнуты благодаря сравнительному изучению мембран из множества разнообразных организмов. Данные, полученные при изучении клеток млекопитающих методом электронной микроскопии, свидетельствуют о наличии развитой сети внутриклеточных мембранных образований. Основные принципы структурной организации всех мембран животной клетки, по сути, одинаковы. Более того, эти принципы распространяются и на мембраны растительных и бактериальных клеток (рис. 1).
Рис. 1. Органеллы эукариотических клеток животных и растений
Любая клетка имеет наружную мембрану – ее называют плазматической. Она играет роль преграды, отделяющей живое содержимое от ее окружения. Она регулирует поступление молекул и ионов в клетку и выход их наружу. Кроме того, в ней находятся различные ферменты, природа которых зависит от особенностей данной клетки. Она содержит специализированные компоненты, участвующие в межклеточных взаимодействиях, в гормональном ответе и системах транспорта через мембрану малых и больших молекул. Плазматическая мембрана чрезвычайно эластична, благодаря чему животные клетки могут изменять форму без разрыва мембран.
1
Большинство растительных клеток в отличие от животных не способны изменять свою форму, так как их мембраны окружены толстой, прочной и малоупругой оболочкой. Ее называют клеточной стенкой. Стенки имеются также у бактерий. Бактериальные клетки имеют довольно простую наружную оболочку, содержащую один или два слоя.
Внутри клетки мембраны могут образовывать органеллы различного назначения. Их внешний вид неодинаков в клетках разного типа. К распространенным внутриклеточным органеллам относятся митохондрии – в них осуществляются окислительные превращения субстратов, завершающиеся образованием АТФ. Независимо от размера и формы каждая митохондрия содержит две мембраны — наружную и внутреннюю. Внутренняя мембрана образует складки в виде перегородок, называемых кристами, и содержит ферменты, участвующие в транспорте электронов и синтезе АТФ. Пространство, ограниченное этой мембраной, носит название матрикса, в нем протекают многие метаболические процессы.
Другие субклеточные органеллы – лизосомы – представляют собой окруженные мембранами органеллы, содержащие набор протеолитических и других деградационных ферментов, которые расщепляют белки, нуклеиновые кислоты, полисахариды и другие соединения. Вещества, захваченные клеткой путем эндоили фагоцитоза доставляются в лизосомы с помощью везикул фагосом.
Фагосомы – это короткоживущие внутриклеточные везикулы, образованные в результате фагоцитоза – процесса поглощения, захвата крупных частиц, комплексов, вплоть до целых клеток, например, бактерий. Этот процесс характерен только для клеток некоторых типов (амебы, макрофаги).
Пероксисомы – это органеллы, которые содержат окислительные ферменты, участвующие в деградации малых молекул, таких как аминокислоты, ксантин. жирные кислоты. Их название связано с присутствием в них каталазы, которая разлагает перекиси, образующиеся в качестве продуктов окисления.
Иной вид внутриклеточных мембран образует так называемую эндоплазматическую сеть – глубокие складки, непосредственно примыкающие к плазматической мембране. Это сложная сеть цистерн и трубочек, которая занимает значительную часть внутреннего объема клетки. На мембранах шероховатого эндоплазматического ретикулума (ЭР) расположены рибосомы. У бактерий, которые не имеют эндоплазматической сети, синтез белка, по-видимому, осуществляется на особых выступах плазматической мембраны. Области ЭР, не содержащие рибосом, называют гладким ЭР. Здесь осуществляется биосинтез стеролов, происходит десатурация (образование двойных связей) жирных кислот.
Мембраны эндоплазматической сети выполняют и другую важную функцию – они обезвреживают вещества, присутствие которых нарушает нормальную работу клетки. Этот процесс называется детоксикацией.
К мембранам эндоплазматической сети примыкает так называемый аппарат
2
Гольджи, также состоящий из ограниченных мембранами пузырьков и цистерн, собранных в стопки. В аппарате Гольджи молекулы белка плотно упаковываются в гранулы – такая упаковка белков происходит перед их секрецией. В аппарате Гольджи происходит также созревание сложных белков, например посттрансляционная модификация гликопротеинов. синтезированных в ЭР и предназначенных для секреции, включения в плазматическую мембрану или доставки в лизосомы. Таким образом, деятельность аппарата Гольджи связана как с построением клеточных элементов, так и с их разрушением.
Оболочка, окружающая клеточное ядро, состоит из двух мембран, наружной и внутренней, разделенных промежутком, называемым перинуклеарным пространством. Эта мембрана происходит из эндоплазматического ретикулума и неразрывно связана с ним. Ядерная мембрана защищает генетическую информацию от вредных внешних воздействий. Наиболее характерными морфологическим признаками ядерной мембраны являются порообразные структуры диаметром около 600 Ǻ. В том месте, где расположены эти структуры, внутренняя и наружная ядерные мембраны соединяются. По краям пор наружная и внутренняя мембраны ядра сливаются в одну общую. Полагают, что поры позволяют комплексам мРНКбелок переходить из ядра в цитоплазму, а регуляторным белкам перемещаться в обратном направлении, из цитоплазмы в ядро. Таким образом, ядерная мембрана контролирует перенос информации между ядром и остальной частью клетки.
Кроме органелл, характерных для большинства клеток, имеются и специализированные мембранные системы, такие как саркоплазматический ретикулум мышечных клеток, миелиновая оболочка периферических нервных волокон,
тилакоидные мембраны хлоропластов и мембраны дисков в палочках сетчатки.
У прокариотичсских организмов также имеются мембраны, хотя и не настолько сложно устроенные, как у эукариотических организмов. Грамположительные бактерии, например, Bacillys subtilis имеют лишь цитоплазматическую мембрану, а грамотрицательные бактерии (Escherichia coli) - еще и наружную оболочку, расположенную поверх тонкой пептидогликановой клеточной стенки. В клетках прокариот обнаружены также некоторые специализированные органеллы (в частности, хроматофоры, содержащие фотосинтезирующий аппарат у пурпурных бактерий).
Хлоропласты – это органеллы, имеющие фотосинтетический аппарат. Они характерны для растений и некоторых микроорганизмов. Наружная оболочка хлоропластов образуется двумя мембранами, а внутренняя область составляет строму.
Все клеточные мембраны отличаются исключительной тонкостью, при этом обладают высокой прочностью на разрыв, устойчивостью и гибкостью, а по электроизоляционным свойствам превосходят многие изоляционные материалы, применяемые в технике. Общая площадь мембран в органах и тканях организма достигает огромных размеров. Печень крысы массой 6 г обладает суммарной
3
площадью клеточных мембран в несколько сотен квадратных метров. В эндоплазматической сети печени на каждый миллиграмм белка приходится 0,5 м2 мембран.
Функции мембран
Несмотря на многообразие типов клеток, их мембраны выполняют общие биологические функции. Прежде всего, они отграничивают живое от неживого. Они организуют внутри клетки компартменты с различными свойствами. С их помощью происходит отделение содержимого компартментов от окружающей их среды. В каждом компартменте мембраны обеспечивают сохранение специфических физико-химических условий. Поэтому по обе стороны мембраны такие условия среды, как кислотность, концентрация растворенных веществ, электрический потенциал, как правило, не одинаковы.
Мембраны участвуют в регуляции метаболических сигналов, которые передаются между наружной и внутренней сторонами. Это может проявляться в виде физического переноса ионов или молекул через мембрану или при помощи конформационных изменений, индуцируемых в мембранных компонентах. Таким образом, мембраны контролируют проникновение в клетку и выход из нее метаболитов. С помощью мембранных рецепторов они реагируют на внешние сигналы и трансформируют их, другими словами, способны классифицировать и избирательно модулировать (усиливать важные и снижать до уровня шумов второстепенные), передавая внутрь клетки существенную информацию.
Мембраны способны обеспечивать образование и поддержание разности потенциалов, а также транспортировать мембранный потенциал вдоль по мембране, позволяя использовать этот специфический вид энергии в разных частях клетки.
Смембранами связано функционирование многих клеточных ферментов. Мембраны оказывают большое влияние на процессы, протекающие внутри клетки, изменяя их активность. Некоторые ферменты активны только тогда, когда они прикреплены к мембране; другие, наоборот, в этом состоянии не проявляют активности и начинают действовать лишь после диссоциации их и выхода в цитоплазму. Одновременно мембранные липиды контролируют взаимодействие между отдельными белками, погруженными в мембранную толщу. Некоторые ферменты образуют своеобразные мембранные ансамбли, которые осуществляют цепь последовательных превращений именно благодаря тому, что их компоненты объединены общностью локализации, т.е. организованы мембраной. Благодаря этому обстоятельству повышается эффективность суммарного процесса.
Сучастием мембран в той или иной степени осуществляется большинство жизненно важных функций. Например, протекают такие процессы, как репликация прокариотической ДНК, биосинтез белков и их секреция, биоэнергетические превращения, а также функционирование систем гормонального ответа. Важная
4
сторона ферментативной деятельности мембран связана с координацией множества химических реакций в клетке. Для этого мембраны объединяют различные ферменты в единый конвейер, в котором каждый фермент действует в строгом соответствии с остальными.
Мембраны участвуют во взаимодействии клеток со средой. Это свойство лежит в основе обеспечения специфики межклеточных контактов и иммунологических ответов.
Большинство мембран, кроме этих общих функций, выполняют и специальные функции. Например, мембраны митохондрий и хлоропластов осуществляют трансформацию энергии. Мембраны клеток, расположенных в стенках кишечника, выполняют функции, связанные с процессами пристеночного пищеварения. Мембраны нервных клеток генерируют электрические импульсы. Некоторые клетки, например палочки сетчатки глаза, имеют высокоспециализированные мембраны, позволяющие выполнять уникальные функции. Мембраны мышечных клеток участвуют в инициации и регуляции сокращения. Клетки органов чувств содержат специализированные мембраны, преобразующие энергию физикохимических стимулов в электрические импульсы и передающие центральной нервной системе информацию о запахах, свете, звуке, изменениях температуры и давления.
Состав мембран
В биомембранах встречаются представители трёх классов веществ: белки, жиры (липиды) и углеводы. По массе белки составляют около 40-60%, остальное преимущественно приходится на липиды. Углеводы чаще входят в гликолипиды и гликопротеины, редко представлены свободными олигосахаридами. Белки чрезвычайно разнообразны, они в значительной степени определяют функциональные свойства и активность мембран (рис.2).
Рис. 2 Схема строения цитоплазматической мембраны
5

Мембранные белки почти не отличаются от цитоплазматических по количеству гидрофобных аминокислот. Однако гидрофобные аминокислоты сгруппированы в мембране в ряд доменов так, что гидрофильных групп полипептидной цепи не хватает для их маскировки. Поэтому такие белки не активны вне гидрофобного окружения мембраны.
Мембранные липиды
В состав мембран эукариотических клеток входят липиды следующих классов:
фосфолипиды, сфинголипиды, гликолипиды и стероид холестерин. Основными липидами мембран животных клеток являются глицерофосфолипиды: фосфатидилхолин и фосфатидилэтаноламин (рис.3 и рис 4).
Рис.3 Фосфолипиды мембраны |
Рис.4 Фософолипиды мембраны. |
В названиях фосфолипидов, потерявших одну из двух ацильных цепей вводится приставка «лизо-». Лизофосфолипиды обнаруживаются в мембранах в небольших количествах – появление этих веществ приводит к нарушению структуры бислоя и лизису клеток.
Наиболее часто встречающиеся фосфолипиды построены по единому плану, их молекулы стерически хорошо соответствуют друг другу. Фосфолипиды очень разнообразны, что связано с различием жирных кислот, входящих в их состав.
Мембраны включают стероиды – спирты со стерановым скелетом. В стероидный ряд мембранных компонентов входят холестерин, ситостерин, тетрахименин. У животных распространен холестерин, в растениях его нет, его заменяет ситостерин. У бактерий стероиды отсутствуют.
6

Рис. 5 Холестерин в мембране
Молекулы холестерина, как и другие липидные молекулы, имеют полярную головку и вытянутую в длину неполярную часть (рис 5). Поэтому они хорошо встраиваются в бислойные липидные структуры, образующие клеточные мембраны. Особенно много холестерина в плазматических мембранах, в печени – это 30% всех липидов мембран.
Холестерин играет роль регулятора, обеспечивающего правильную упаковку липидной части мембраны, необходимую для нормальной работы. Это означает, что если мембрана слишком ригидна и существует опасность «застывания» жирнокислотных цепей, холестерин вызывает ее разжижение, поскольку цепи в его присутствии становятся более подвижными. Если мембрана слишком «жидкая», то холестерин ее уплотняет.
Природа уплотняющего действия холестерина в следующем. Обычно углеводородные хвосты фосфолипидов располагаются под некоторым углом к плоскости мембраны, а не перпендикулярно. В присутствии же холестерина наклон хвостов становится меньше. Каждая молекула лецитина занимает в присутствии холестерина меньшую площадь на поверхности ме6мбраны, в результате чего мембрана уплотняется.
Пространственная конфигурация жирных кислот
Насыщенные ж.к. могут принимать множество конфигураций вследствие высокой свободы вращения вокруг одиночных С-С связей. Энергетически выгодна транс-конформация.
7

Ненасыщенные ж.к. имеют жесткую структуру, т.к. вращение вокруг двойных связей невозможно. Ненасыщенные ж.к. почти всегда содержат двойные связи в цис-конформации (рис 6). Цис-конформация двойной связи обуславливает изгиб двойной связи под углом около 30о, что вызывает локальные возмущения бислоя. В области локализации двойных цис-связей образуются изгибы, т.н. гош-форма.
Рис. 6 Пространственная конформация жирных кислот
При повышении температуры тепловая подвижность жирнокислотных цепей приводит к спонтанному возникновению изгибов. Если изгибы, соответствующие гош-конформации, появляются на близлежащих участках жирнокислотной цепи, эта область принимает вид петли или полости, т.н. кинк. В результате взаимопревращений транс- и гош-конформаций (транс-гош-переход) кинки могут скользить вдоль цепи, обеспечивая перемещение их содержимого поперек мембраны. Т.о. может происходить диффузия захваченной ими воды через гидрофобный бислой.
Фосфолипиды – структурная основа бислоя
Природные фосфолипиды – выраженные амфифилы. Величина ККМ у них очень маленькая, т.е. уже при низких концентрациях при комнатной температуре
8
они образуют упорядоченные мицеллы, однослойные или многослойные агрегаты –
липосомы.
Бислойная структура стабилизируется гидрофобными взаимодействиями в области ацильных связей и полярными взаимодействиями на границе раздела водной и липидной фаз. Основу полярных взаимодействий обеспечивают силы: ионные, диполь-дипольные, водородные и вандерваальсовские. Ацильные гидрофобные хвосты прячутся в середине в силу термодинамической выгодности. Гидрофобные силы – это способность аполярных групп к тесному контакту в водных средах, которое обеспечивает вытеснение воды из образуемых агрегатов.
Головки несут заряд либо минус, либо минус и плюс одновременно, нейтрализующие друг друга. Нейтральный тип преобладает в большинстве клеточных мембран.
Трансмембранная асимметрия липидов
Трансмембранная асимметрия – это различие липидного и белкового состава внешней и внутренней поверхности. Асимметрия липидов возникает, главным образом, оттого, что в случае замкнутого мембранного бислоя липиды с более объемными головками стремятся в наружный слой. Поэтому появляются различия в заряде фосфолипидов и разной гидратации внутренней и внешней поверхности.
Кроме того, различия в ионном составе вызывают разные изгибы мембраны, т.е. асимметрию бислоя.
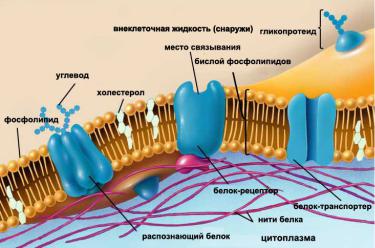
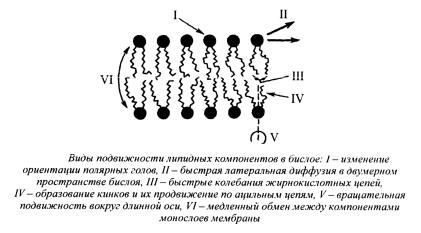
Рис. 7 Виды подвижности липидов в мембране
Подвижность компонентов бислоя
Молекулы фосфолипидов способны к нескольким видам подвижности в бислое (рис 7 и 8):
1.изменение ориентации полярных головок,
2.латеральное движение,
9
3.колебания ацильных цепей,
4.образование кинков и их движение в поперечном направлении,
5.ротационная подвижность (вращение вокруг длинной оси),
6.переход с одной стороны слоя на другую (флип-флоп),
7.выход из бислоя.
Рис. 8 Виды подвижности липидов в мембране
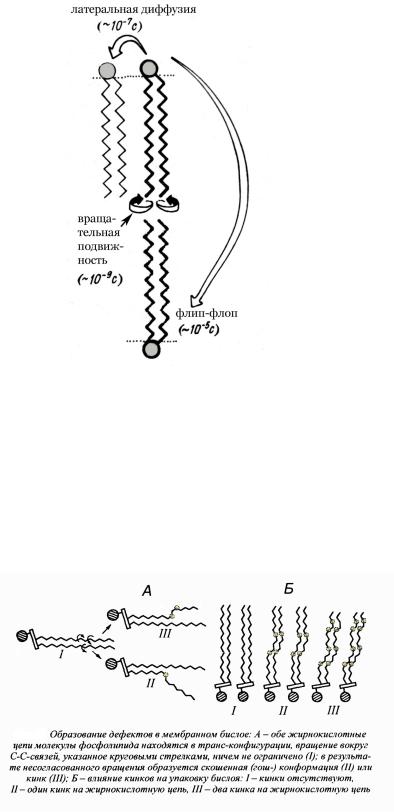
Дефектные зоны
Подвижность липидных компонентов в бислое нарушает гомогенную упаковку и приводит к образованию различных дефектов. Высокая скорость вращения жирнокислотных радикалов вокруг С-С-связей, наличие цис-изомеров ненасыщенных ж.к. делают бислой достаточно рыхлым (рис 9).
Рис. 9. Дефекты липидного бислоя
10