

Химическое строение и молекулярная структура
Целлюлоза представляет собой природное высокомолекулярное органическое соединение, состоящее из большого числа повторяющихся элементарных звеньев [С6Н7О2(ОН)3]n.
Считается, что биосинтез целлюлозы в растительных клетках и тканях протекает по следующему механизму:
гуанозинтрифосфат + -D-глюкоза-1-фосфат ↔ гуанозиндифосфат-D-глюкоза + пирофосфат
n (гуанозиндифосфат-D-глюкоза) + акцептор → акцептор[-(1→4)-D-глюкоза]n + n гуанозиндифосфат
где роль акцептора выполняет фермент, переносящий активированные D-глюкозные остатки на растущую полисахаридную цепь.
Целлюлоза – гомополисахарид.Макромолекулы целлюлозы представляют собой линейные неразветвленные цепи, построенные из большого числа остатков -D-глюкопиранозы (ангидроглюкозных звеньев), соединенных между собой -(1→4)-гликозидными (ацетальными) связями (рис.4).
|
|
|
Рис.4. Структурная формула макромолекулы целлюлозы, n – степень полимеризации. |

Ацетальная (гликозидная) связь – связь, образованная в результате взаимодействия альдегидной группы одного элементарного звена макромолекулы со спиртовой гидроксильной группой другого звена. (Нумерация углеродных атомов в элементарном звене макромолекулы целлюлозы показана на рис.4.) Цифры 1→4 соответствуют нумерации углеродных атомов глюкопиранозного цикла, участвующих в образовании гликозидной связи. Важной характеристикой строения макромолекулярной цепи целлюлозы (да и всех полисахаридов) является не только направление гликозидной связи, но и ее конфигурация. Гликозидная связь в макромолекуле целлюлозы имеет -конфигурацию (это означает, что одно глюкопиранозное кольцо повернуто вокруг своей оси на ~180 относительно соседнего, т.е. имеет противоположную пространственную ориентацию по сравнению с конфигурацией асимметричного углеродного атома С5 в молекуле глюкозы).
Наличие асимметричного атома углерода придает глюкопиранозному циклу макромолекулы целлюлозы хиральность, то есть неэквивалентность зеркальному изображению. Это приводит к возникновению оптической активности звена и макромолекулы в целом. Глюкопиранозные циклы могут иметь две конформации типа кресла и шесть конформаций типа ванны (рис.5).
На основании главным образом квантово-химических и конформационных расчетов была показана предпочтительность конформации кресла С1. Энергия перехода из конформации кресла в конформацию ванны, рассчитанная с использованием модели атом-атомных взаимодействий, составляет 54 кДж/моль. Для обратного перехода значение этой величины - 26 кДж/моль. Это свидетельствует о большей устойчивости и меньшей напряженности конформации кресла. Гидроксильные и гидрометильная группы в конформации С1 расположены экваториально eq, т.е. располагаются приблизительно в плоскости, образуемой вторым, третьим и пятым атомами углерода и атомом кислорода пиранозного цикла.
|
|
|
|
Рис.5. Возможные конформации глюкопиранозных циклов. | |



Наличие в глюкопиранозных циклах макромолекул целлюлозы боковой СН2ОН-группы обуславливает возможность появления поворотных изомеров. С помощью критерия минимума энергии выделят три заторможенные конформации, соответствующие наиболее вероятным изомерам при вращении группы –СН2ОН вокруг связи С5 – С6. Ниже приведены эти три конформации для связи С6 – О6:
- гош по отношению к С5 – О5 и транс по отношению к С4 – С5 (gt);
- гош по отношению к С5 – О5 и гош по отношению к С4 – С5 (gg);
- транс по отношению к С5 – О5 и гош по отношению к С4 – С5 (tg).
Конформации gt-, gg- и tg- схематически показаны на рис.6. Структурные исследования методом поляризованной ИК-спектроскопии показали, что наиболее предпочтительна gt-конформация.
|
(а) |
(б) |
|
|
|
|
Рис.6. Возможные конформации группы –СН2ОН: пиранозный цикл (а), боковые проекции (б). | |


Элементарной единицей макромолекулы целлюлозы является целлобиозный остаток, состоящий их двух глюкопиранозных циклов. Обычно конформацию целлобиозного остатка изображают так, как это показано на рис.7. Там же указаны внутримолекулярные водородные связи (Н-связи) О2…(Н)О6 и О5…(Н)О3. Эти связи фиксируют взаимное положение соседних звеньев. Расчетная длина связи О2…(Н)О6 составляет 3.83 Å, а О5…(Н)О3 2.68 Å.
|
|
|
Рис.7. Конформация целлобиозного остатка. |
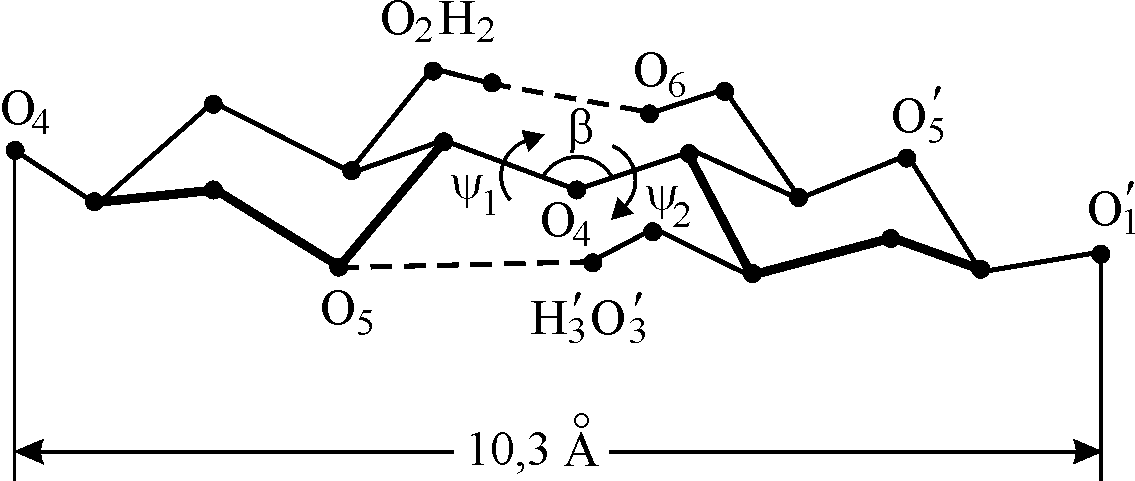
Анализ оптимизированных конформационных карт димерного фрагмента целлюлозной цепи показал, что наиболее устойчив конформер с ψ1 = – 210º и ψ2 = – 150º. Это приводит к «закрученности макромолекулы». Теоретический расчет возможных ориентаций глюкопиранозных циклов с помощью критерия допустимых контактов привел к выводу о спиральной конформации целлюлозной цепи с шагом 0.515 нм и с числом мономерных звеньев два-три на виток. Таким образом, в высокоупорядоченных областях макромолекулы целлюлозы будут иметь жесткие спиральные конформации, стабилизированными внутримолекулярными Н-связями.
Кроме внутримолекулярных Н-связей в целлюлозе имеются и межмолекулярные связи типа О6…(Н)О3 и О6…(Н)О2 (рис.8).
|
|
Рис.8. Система внутримолекулярных и межмолекулярных Н-связей в целлюлозе. |

Возможно также образование межмолекулярных Н-связей типа О5…(Н)О3 и О2…(Н)О6.
Элементарная ячейка пространственно-структурной организации целлюлозы показана на рис.9. Группы –СН2ОН связаны межмолекулярными водородными связями с глюкозидными атомами кислорода и атомами кислорода глюкопиранозного кольца почти параллельными «плоскостями». Цепь, расположенная в центре элементарной ячейки сдвинута по оси макромолекулы примерно на половину элементарного звена. При этом кислородный атом пиранозного кольца попадает в энергетически выгодное положение по отношению к оксиметильным группам угловых цепей. В приведенной на рис.9 схеме оксиметильные группы связаны с мостичными атомами кислорода.
|
(а) |
Рис.9. Схематическое изображение внутри- и межмолекулярных Н-связей в элементарной ячейке целлюлозы: со стороны плоскости ав (а), вдоль оси волокна (б). |
|
| |
|
(б) | |
|
|


Наличие в целлюлозы системы внутри- и межмолекулярных Н-связей приводит к тому, что несколько целлюлозных цепей довольно прочно связываются друг с другом посредством водородных связей между гидроксильными группами и межмолекулярных сил с малой энергией взаимодействия (вандерваальсовыми связями и т.п.) таким образом, что образуется жесткая пространственная, так называемая «вторичная» структура полимера.