

Подготовка у универсиаде 2012 / Генетика (Жимулев) / 6ver7
.pdfСтруктура и организация генома |
|
|
|
|
|
Глава 6 |
|||||
|
|
|
|||||||||
Глава 6.Cтруктура и |
|
оболочках,а70%метки32Р-винфицированных |
|||||||||
организация генома |
|
бактериях. Фаги-потомки получили только |
|||||||||
|
около1%исходногобелка,меченого35S,однако |
||||||||||
|
|
|
|
||||||||
6.1. Ðîëü ÄÍÊ â |
|
онижеобнаружилиоколо30%метки32Ð. |
|||||||||
|
Результаты этого эксперимента прямо |
||||||||||
наследственности |
|
||||||||||
|
показали, что ДНК родительских фагов |
||||||||||
|
|
|
|
||||||||
|
По-видимому,самуюпервуюгипотезуо |
проникает в бактерии и затем становится |
|||||||||
физико-химическойприродегеновихромосом |
составляющей развившихся новых фаговых |
||||||||||
предложил Н.К. Кольцов в 1927, а в более |
частиц. |
|
|
|
|
|
|
||||
развернутой форме – в 1935 |
|
|
|
|
|
|
|
|
|||
Дополнение 6.1 |
|
|
|
|
|
|
|
||||
году(Дополнение6.1) (Детали |
Н.К. Кольцов предположил, что хромосомы представляют |
||||||||||
см. также в статьях В.А. |
|||||||||||
собой огромные молекулы белков или пучки таких молекул. О |
|||||||||||
Ратнера 1998а, б). |
существовании в клетках других длинных молекул, состоящих из |
||||||||||
|
Как уже упоминалось в |
гетерогенных мономеров, тогда еще не знали. ДНК, или как ее тогда |
|||||||||
Разделе 5.11, в результате |
называли – тимонуклеиновая кислота – считалась, как пишет Э. |
||||||||||
изучения |
явления |
Шредингер в своей знаменитой книге «Что такое жизнь с точки зрения |
|||||||||
физика?» «сравнительно простым органическим соединением, |
|||||||||||
трансформацииубактерийбыло |
которому было бы странно приписывать роль носителя |
||||||||||
впервыепоказано,чтоименно |
наследственных свойств». Вот что пишет сам Н.К. Кольцов о |
||||||||||
ÄÍÊ |
может |
служить |
формировании хромосом (генонем) и генов: |
|
|
|
|
|
|||
генетическим материалом. В |
«Первоначально, когда у простейших организмов впервые |
||||||||||
1952 году были получены |
слагались генонемные молекулы, они были представлены |
||||||||||
новые доказательства этого в |
однообразными более или менее длинными цепями из одинаковых |
||||||||||
звеньев, вроде кератина или серицина. Каждое звено состояло из |
|||||||||||
экспериментах другого типа. |
немногих простых радикалов. При дальнейшей эволюции организма |
||||||||||
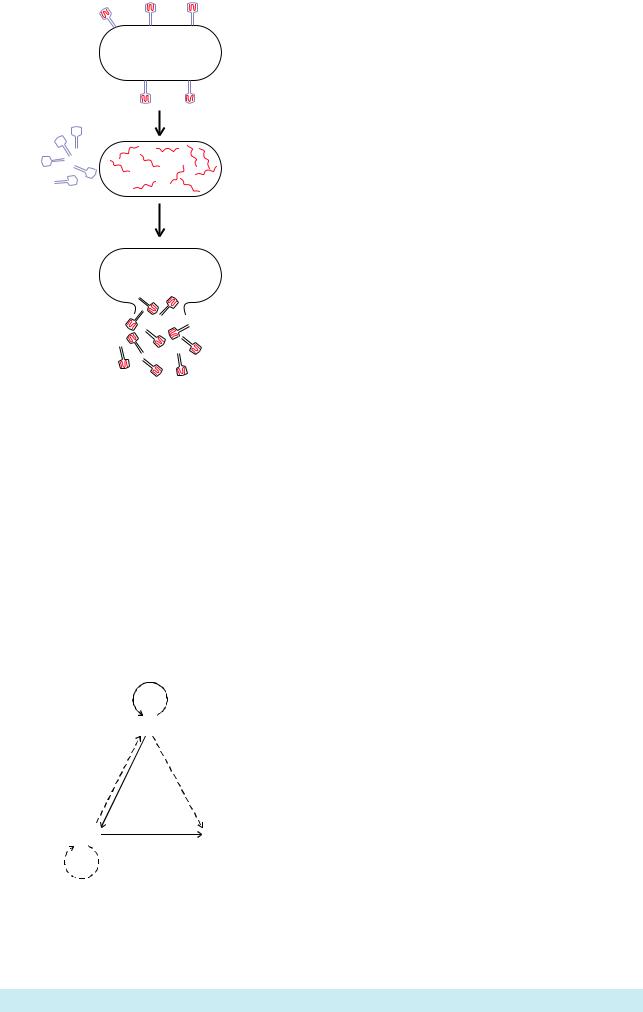
Как известно, фаг Т2 является |
эти молекулы постепенно усложнялись путем присоединения к |
||||||||||
вирусом, инфицирующим |
некоторым звеньям боковых радикалов, получающих значение генов. |
||||||||||
бактерию E. coli. Фаговые |
Мало-помалу число этих боковых цепей, размещенных в |
||||||||||
частицы абсорбируются на |
определенных пунктах генонемы, увеличивалось и самые радикалы |
||||||||||
наружнойповерхностиклетки, |
|
âñå |
более |
|
усложнялись. |
||||||
|
Микроскопическая |
картина |
|||||||||
|
|
|
|
||||||||
ихматериалпроникаетвнутрьи |
|
хромосом в слюнных железах |
|||||||||
примерно через 20 минут |
|
дрозофилы представляет картину |
|||||||||
бактерия |
лизируется, |
|
óæå |
|
очень |
|
высокой |
||||
освобождая |
большое |
|
дифференцировки генонем. Если |
||||||||
|
признать, что поперечные диски |
||||||||||
количество фаговых частиц - |
|
||||||||||
|
соответствуют генам, то здесь мы |
||||||||||
потомков. В |
1952 ãîäó |
|
|||||||||
|
должны поместить именно боковые |
||||||||||
АльфредХершииМартаЧейз |
|
радикалы или цепи радикалов, |
|||||||||
инфицировалибактерийфагами |
|
которые |
адсорбируют |
ÿðêî |
|||||||
Т2, которые были мечены |
|
окрашенный хроматин. В таком |
|||||||||
|
случае неокрашиваемые сегменты, |
||||||||||
радиоактивнымисоединениями: |
|
||||||||||
|
в которых |
ìû |
различаем |
||||||||
ДНК-спомощью32Р,белковая |
|
||||||||||
|
продольные нити, придется признать |
||||||||||
|
|
|
|
||||||||
частьфага-35S(Рис.6.1).После |
|
основными |
|
цепями, |
íå |
||||||
инфекции бактерий фагами, с |
|
осложненными |
|
сложными |
|||||||
помощьюцентрифугирования |
|
боковыми придатками. Но при |
|||||||||
удалосьвыделитьдвефракции: |
|
дальнейшей дифференцировке и |
|||||||||
|
ñþäà |
могут |
присоединяться |
||||||||
пустые белковые оболочки |
|
||||||||||
|
боковые радикалы – новые гены, а |
||||||||||
ôàãà |
è |
бактерий, |
|
||||||||
|
с другой стороны, уже имеющиеся |
||||||||||
инфицированныхфаговойДНК. |
|
боковые |
радикалы |
могут |
|||||||
Оказалось, что 80% метки 35S |
|
осложняться |
è |
упрощаться в |
|||||||
осталась в пустых фаговых |
|
мутационном процессе ”(Кольцов, |
|||||||||
|
1935, ñòð. 119.). |
|
|
|
|
||||||
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
107
Глава 6 |
|
|
|
Структура и организация генома |
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Рисунок 6.1 |
|
|
|
Дополнение 6.2 |
|
|
|
|
|
|
В 1969 году Альфред Д. Херши (A.D. |
||
|
|
|
|
Hershey) получил Нобелевскую премию за |
||
|
|
|
|
открытие генетической структуры вирусов. |
||
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
Дополнение 6.3 |
|
|
|
|
|
|
За исследования нуклеотидов и |
||
|
Встряхивание |
|
|
нуклеозидов Нобелевскую премию в 1957 |
||
|
|
|
|
году получил Александр Тодд (A. Todd). |
||
|
|
|
|
РНК,РНК→ белок,РНК→ РНК.2.Процессы, |
||
|
|
|
которыенебылиэкспериментальновыявлены |
|||
|
|
|
и с теоретической точки зрения не казались |
|||
|
Лизис |
|
строго необходимыми: РНК → |
ÄÍÊ, ÄÍÊ → |
||
|
|
|
белок. 3. Невозможные переносы: белок → |
|||
|
|
|
белок, белок → |
РНК, белок → |
ДНК. Таким |
|
|
|
|
образом,информациявовсехслучаяхвклетке |
|||
|
|
|
переноситсяоднонаправленнопоцепи:ДНК→ |
|||
|
|
|
РНК → белок. Белок не может служить |
|||
|
|
|
матрицейдлясинтезаДНКилиРНК,поскольку |
|||
|
|
|
у молекул белка нет свойства |
|||
Генетическим материалом фага T2 является |
|
комплементарности отдельных частей |
||||
ДНК. Белки фага мечены !#S (синий цвет), |
|
молекулы, что бы позволяло использовать е¸ |
||||
ÄÍÊ - ! P (красный цвет). |
|
какматрицу. |
|
|
||
СовременныепредставленияоролиДНК |
|
|
|
|
||
впередаченаследственнойинформациилучше |
|
6.2. Структура ДНК |
|
|||
всего отражает |
“Центральная догма |
|
Генетическая информация в молекуле |
|||
молекулярнойбиологии”,сформулированнаяФ. |
ДНК записана в виде последовательности |
|||||
Крикомв1956идоработаннаяв1970годах(Рис. |
нуклеотидных остатков, которые содержат |
|||||
6.2). |
|
|
одноизчетырехазотистыхоснований:аденин |
|||
Автор предложил разделить все виды |
(А), гуанин (G), цитозин (С) и тимин (Т) (Рис. |
|||||
переносабиологическойинформациивклетке |
6.3). |
|
|
|||
на три группы: 1. Процессы, существование |
|
|
Азотистые основания делятся на два |
|||
которых уже показано: ДНК → ДНК, ДНК → |
типа: пиримидиновые и |
пуриновые. |
||||
|
|
|
Пиримидинысостоятизшестичленногокольца, |
|||
Рисунок 6.2 |
|
|
||||
|
|
аупуриновподвакольца:одно–пятичленное |
||||
|
|
|
||||
|
|
|
ивторое–шестичленное.Каждаянуклеиновая |
|||
|
ÄÍÊ |
|
кислота синтезируется из оснований только |
|||
|
|
|
|
|
|
|
|
|
|
четырехтипов.Одниитежепурины(аденини |
|||
|
|
|
гуанин) входят в состав и ДНК И РНК. Два |
|||
|
|
|
пиримидина, входящие в состав ДНК, - это |
|||
|
|
|
цитозин и тимин, а в РНК вместо тимина |
|||
ÐÍÊ |
Белок |
|
находитсяурацил.Тиминотличаетсяотурацила |
|||
|
только наличием метильной группы в пятом |
|||||
|
|
|
||||
|
|
|
положении |
пиримидинового кольца. |
||
“Центральнаядогма молекулярнойбиологии”. |
|
Соединения,состоящиеизостатковазотистого |
||||
|
основания |
и углевода |
рибозы или |
|||
Сплошные стрелки показывают обычный |
|
|||||
|
дезоксирибозы называются нуклеозидами. |
|||||
путь переноса генетической информации, |
|
|||||
|
Нижеприведенаноменклатураназванийвсех |
|||||
пунктирной - более редкие пути, также |
|
|||||
|
этихсоединений: |
|
||||
существующие в природе (Из: Crick, 1970). |
|
|
||||
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
108

Структура и организация генома |
|
|
|
|
|
Глава 6 |
|||||||||
Рисунок 6.3 |
|
|
|
|
|
|
|
|
|
Рисунок 6.4 |
|
|
|
||
|
|
|
O |
|
|
|
|
|
|
|
5’ |
3’ |
|
||
|
|
N |
|
NH |
|
|
|
|
|
|
|
|
|
0.34 íì |
|
5’ |
|
|
G |
|
|
|
|
|
|
|
|
|
|||
CH2 |
|
N |
N |
NH2 |
|
|
|
|
|
|
Малая |
|
|
|
|
O |
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
NH2 |
|
|
|
|
|
бороздка |
|
|
|
|||
P |
O |
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
||||
H |
H |
|
|
|
N |
|
|
|
|
|
A |
|
|
|
|
H |
|
H |
|
C |
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
H O |
CH2 |
N |
O |
O |
|
|
|
|
G |
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
P |
|
H |
O |
H3C |
|
|
|
|
|
T |
|
|
|
|
O |
|
|
H |
|
T NH |
|
|
|
|
C |
|
|
|||
|
|
H |
H |
|
|
|
|
|
|
|
|||||
|
|
|
|
H O |
CH2 |
N |
O |
|
|
|
|
|
|
|
|
|
|
|
|
|
O |
|
|
NH2 |
Большая |
|
3.4 íì |
||||
|
|
|
|
P |
|
|
|
|
|
бороздка |
|
||||
|
|
|
|
H |
H |
|
N |
|
|
|
|
||||
|
|
|
|
|
|
|
N |
|
|
||||||
|
|
|
O |
|
|
|
|
|
|
|
|||||
|
|
|
|
H |
H |
|
|
A |
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
N |
|
|
|
|
|
||||
|
|
|
|
|
|
H O |
CH2 |
N |
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
P |
O |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
H |
H |
|
|
|
|
|
|
||
|
|
|
|
|
O |
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
H |
H |
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
3’ |
H |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
P |
|
|
|
|
|
|
||
|
|
|
|
|
|
|
O |
|
|
|
|
|
|
|
|
Фрагмент одной цепи ДНК. Пуриновые |
|
|
|
|
|||||||||||
основания аденин (А) и гуанин (G) и |
|
|
|
|
|||||||||||
пиримидиновые основания тимин (Т) и |
|
5’ |
3’ |
|
|||||||||||
цитозин (С) прикреплены к полимерному |
|
2.0 íì |
|
|
|||||||||||
остову, состоящему из чередующихся остатков |
Модель структуры ДНК по Уотсону и Крику |
||||||||||||||
фосфата (Р) и сахара дезоксирибозы. |
|
|
(Из: Фаворова, 1996, стр. 13). |
|
|||||||||||
Основания |
Нуклеозиды Нуклеотиды |
|
Дополнение 6.4 |
|
|
||||||||||
|
За открытие структуры нуклеиновых |
||||||||||||||
--------------------------------------------------- |
|||||||||||||||
кислот Френсис Крик, Джеймс Уотсон и Морис |
|||||||||||||||
Аденин (A) |
Аденозин |
Адениловая |
|
||||||||||||
|
Уилкинс (F.H.C. Crick, J.D. Watson, M.H.F. |
||||||||||||||
|
|
|
|
|
|
кислота |
|
|
|||||||
|
|
|
|
|
|
|
|
Wilkins) в 1962 году были награждены |
|||||||
|
|
|
|
|
|
(AMP èëè dAMP) |
|||||||||
|
|
|
|
|
|
Нобелевской премией. |
|
|
|||||||
Гуанин (G) |
Гуанозин |
Гуаниловая |
|
|
|
|
|||||||||
|
|
Каждая |
öåïü |
содержит |
|||||||||||
|
|
|
|
|
|
кислота |
|
|
|||||||
|
|
|
|
|
|
|
|
последовательность нуклеотидов, строго |
|||||||
|
|
|
|
|
|
(GMP èëè dGMP) |
|||||||||
|
|
|
|
|
|
|
|
|
|
||||||
Цитозин (C) |
Цитидин |
Цитидиловая |
|
соответствующуюпоследовательностидругой |
|||||||||||
|
|
|
|
|
|
кислота |
|
|
цепи.Этосоответствиедостигаетсяналичием |
||||||
|
|
|
|
|
|
(CMP èëè dCMP) |
водородных связей между направленными |
||||||||
Тимин(T) |
|
Тимидин |
Тимидиловая |
навстречу друг другу основаниями двух |
|||||||||||
|
|
|
|
|
|
кислота (dTMP) |
цепей: G и С или А и Т. Таким образом, цепи |
||||||||
Урацил (U) |
Уридин |
|
Уридиловая |
|
комплементарны. Поскольку цепи имеют |
||||||||||
|
|
|
|
|
|
кислота (UMP) |
противоположную направленность в |
||||||||
Модель ДНК в форме регулярной |
расположении 5' и 3' свободных концов в |
||||||||||||||
двойной спирали (Рис. 6.4) была предложена |
молекуле |
пентозы, |
èõ |
называют |
|||||||||||
Дж. Д. Уотсоном и Ф. Криком в 1953 году |
антипараллельными. |
|
|
||||||||||||
(Ðèñ. 6.5). |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Двеспиральныеполинуклеотидныецепи |
6.3. Репликация ДНК |
|
|||||||||||||
закручены вправо вокруг общей оси. |
|
|
|
|
|||||||||||
Пуриновые остатки заштрихованы. Против |
6.3.1. Механизмы репликации |
||||||||||||||
каждого |
èç |
íèõ |
находится |
остаток |
УотсониКрикужевовторойсвоейработе |
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|||||
пиримидинового основания другой цепи. На |
1953годапредположиливозможныймеханизм |
||||||||||||||
схеме показаны размеры спирали, наличие |
копированиянаследственногоматериала.Легко |
||||||||||||||
большой и малой бороздок и представить,чтоцепимолекулыДНКрасходятся |
|||||||||||||||
антипараллельностьдвухцепейДНК. |
|
|
икаждаяизнихстановитсяматрицей,накоторой |
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|||||
109

Глава 6 |
|
|
|
|
|
|
Структура и организация генома |
||||
Рисунок 6.5 |
|
|
|
|
|
|
однаизних“консервативная”, |
||||
|
|
|
|
|
|
|
другая-“дисперсионная”(Рис. |
||||
|
|
|
|
|
|
|
6.7).Доказалисуществование |
||||
|
|
|
|
|
|
|
полуконсервативной модели |
||||
|
|
|
|
|
|
|
М.МезелсониФ.Стальв1958 |
||||
|
|
|
|
|
|
|
году. Авторы выращивали |
||||
|
|
|
|
|
|
|
бактерии E. coli несколько |
||||
|
|
|
|
|
|
|
поколений на минимальной |
||||
|
|
|
|
|
|
|
среде, |
â |
которой |
||
|
|
|
|
|
|
|
единственным источником |
||||
|
|
|
|
|
|
|
азота был |
15NH Cl (хлорид |
|||
|
|
|
|
|
|
|
|
|
|
4 |
|
|
|
|
|
|
|
|
аммония).Вэтомсоединении |
||||
|
|
|
|
|
|
|
нормальный изотоп 14N, áûë |
||||
|
|
|
|
|
|
|
заменен на 15N. В результате |
||||
|
|
|
|
|
|
|
все клеточные компоненты |
||||
|
|
|
|
|
|
|
бактерий, включая пурины и |
||||
|
|
|
|
|
|
|
пиримидинывмолекулахДНК |
||||
|
|
|
|
|
|
|
содержалиболеетяжелыйазот |
||||
Д. Уотсон (род. 1928) и Ф. Крик (род. 1916) у стереомодели 15 |
|
|
|
||||||||
молекулы ДНК. |
|
|
|
|
N.Затемклеткипереносили |
||||||
|
|
|
|
насреду,содержащуюизотоп |
|||||||
|
|
|
|
|
|
|
|||||
синтезируетсяноваякомплементарнаяцепь(Рис. |
14N. Через 1 или 2 поколения |
||||||||||
выделялиДНКицентрифугироваливградиенте |
|||||||||||
6.6). В результате образуются две дочерние |
|||||||||||
двуспиральныемолекулыДНК,неотличимыеот СsСl.Фракции,содержащиелегкиеилитяжелые |
|||||||||||
родительскоймолекулы. |
|
|
цепи, а так же гибридные 15N/14N, легко |
||||||||
|
|
отделялись одна от другой (Рис. 6.8). ДНК, |
|||||||||
КаждаямолекулаДНКсостоитизодной |
|||||||||||
цепиисходнойродительскоймолекулыиодной выделеннаяизбактерийпервогопоколения,дает |
|||||||||||
вновьсинтезированнойцепи.Такоймеханизм прицентрифугированииоднуполосу,состоящую |
|||||||||||
копированияназываетсяполуконсервативным.В |
из гибридных 15N/14N цепей, а во втором |
||||||||||
то же время обсуждались две другие модели, |
поколении–двеполосы:из14N/14Nè15N/14N,÷òî |
||||||||||
Рисунок 6.6 |
|
|
|
|
|
|
свидетельствует о полуконсервативной схеме |
||||
|
|
|
|
|
|
репликации(см.Рис.6.7). |
|
|
|||
|
|
|
|
|
|
|
|
|
|||
|
|
T |
|
A |
|
|
В1957годуАртурКорнбергобнаружилу |
||||
|
|
|
|
|
|
|
|
|
|
||
|
|
C |
G |
|
|
бактерии E. coli фермент, катализирующий |
|||||
Родительская |
|
|
|
|
|
Родительская |
процессполимеризацииДНКизнуклеотидов- |
||||
öåïü |
|
|
|
A |
T |
öåïü |
ДНК-полимеразу. |
Открытие Корнберга |
|||
|
|
|
|
|
|
|
|||||
|
|
|
|
G |
C |
|
показало, что в основе удвоения молекул |
||||
|
|
|
|
|
|
|
|
|
|||
|
|
G |
|
|
C |
|
ДНК лежат обычные биохимические |
||||
|
A |
|
Одиночные |
T |
реакции. По современным представлениям, |
||||||
|
|
родительские |
в репликации ДНК у прокариот выделяют |
||||||||
|
|
|
|||||||||
T |
A |
|
|
öåïè |
|
||||||
|
|
|
T |
|
следующие этапы: |
|
|
|
|||
|
|
|
|
|
A |
|
|
|
|||
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
||
C |
G |
|
|
|
C |
|
1. Релаксация суперспирализованной ДНК. |
||||
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
G |
|
|
|
|
|
A |
|
|
|
|
|
|
Этот процесс катализируется ферментом |
||||
|
T |
|
|
A |
T |
топоизомеразой. |
|
|
|
||
|
|
|
|
|
|
|
|
||||
G |
|
C |
|
|
G |
C |
2. Денатурация двойной спирали ДНК. |
||||
|
|
|
|
|
|
|
Поскольку синтез ДНК происходит на |
||||
|
|
Дочерние цепи |
|
одноцепочечной матрице, ему должно |
|||||||
Схема полуконсервативной репликации ДНК |
предшествовать обязательное разделение |
||||||||||
|
|
|
|
||||||||
(Èç:Lewin,1994,ñòð.95). |
|
|
двухцепейДНК.Участокначаларасхождения |
||||||||
|
|
|
|
|
|
||||||
110 |
|
|
|
|
|
|
|
|
|
|
|

Структура и организация генома |
Глава 6 |
|
Рисунок 6.7 |
|
|
à |
á |
â |
Родительские
молекулы
Первое
поколение
Второе
поколение
Модели репликации ДНК. а - полуконсервативная, б - консервативная, в - дисперсионная. Родительские цепи изображены в виде красных лент, вновь синтезированные показаны синим цветом.
цепейназываетсярепликационнойвилкойиз- |
|
|
Дополнение 6.5 |
|
|
|||
|
|
В 1959 году Артуру Корнбергу (Arthur |
||||||
за характерной Y-образной формы (Рис. 6.9). |
|
|
||||||
|
|
Kornberg) была присуждена Нобелевская |
||||||
Именно в этой репликационной вилке |
|
|
||||||
|
|
премия за открытие механизма биосинтеза |
||||||
ДНК-полимеразы синтезируют дочерние |
|
|
||||||
|
|
ÄÍÊ. |
|
|
||||
|
|
|
|
|
|
|
|
|
Рисунок 6.8 |
|
|
|
|
|
|
|
|
|
ÄÍÊ â |
|
|
|
|
|
|
|
Культуры |
градиенте |
|
|
|
|
|
Ôîòî |
|
E. coli |
CsCl |
|
|
|
|
Состав ДНК |
|
фракций |
Поколение 0 |
|
|
|
|
|
Тяжелая ДНК |
||
Среда |
|
|
|
|
|
|||
содержит |
|
|
|
|
|
(15N/15N) |
|
|
15N |
|
|
|
|
|
|
|
|
Поколение 1 |
|
|
|
|
|
Тяжелая/легкая |
||
Ðîñò |
|
|
|
|
|
|||
культур на |
|
|
|
|
|
ÄÍÊ (15N/14N) |
||
среде с 14N |
|
|
|
|
|
|
|
|
Поколение 2 |
Легкая |
Тяжелая/легкая |
||||||
Ðîñò |
ÄÍÊ |
|
||||||
|
15 |
14 |
N) |
|||||
культур на |
|
14 |
|
14 |
|
ÄÍÊ ( N/ |
|
|
( |
N/ |
N) |
|
|
||||
среде с 14N |
|
|
|
|
||||
Поколение 3
Рост культур на среде с 14N
14N/14N 14N/14N 14N/14N 15N/14N
Схема опытов Мезелсона и Сталя, доказывающих полуконсервативную модель репликации ДНК (Из: Russell, 1998, p. 346).
111

Глава 6 Структура и организация генома
Рисунок 6.9 |
|
|
|
|
|
|
|
направлении, с заменой одной из пар |
||||||
|
|
|
Точка начала |
|
|
|
нуклеотидов. Всего у B. subtilis на OriC |
|||||||
|
|
|
|
|
|
|
|
|
|
|
||||
Родительская ДНК |
репликации |
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
расположено 15 ДНК-боксов. Область OriC |
||||||||
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
очень консервативна: ДНК-боксы сходного |
|||||
|
|
|
|
|
|
|
|
|
состава имеются в соответствующем месте |
|||||
|
|
|
Точка начала |
|
|
|
хромосомы и других бактерий (хотя у |
|||||||
|
|
|
репликации |
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
||||
Репликационный глазок |
|
|
|
|
|
|
Mycoplasma genitalium, несмотря на наличие |
|||||||
|
|
|
|
|
|
|||||||||
|
|
|
|
|
|
общих для всех бактерий ферментов |
||||||||
с одной вилкой |
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
репликации, ДНК-боксов найдено не было). |
|||||
|
|
|
|
|
|
|
|
|
С данными |
последовательностями |
||||
|
|
|
|
|
|
|
|
|
происходит связывание белка DNAA, |
|||||
|
|
|
|
|
|
|
|
|
продукта гена dnaA; это событие служит |
|||||
|
|
|
|
|
Репликационная |
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
||||
Репликационный глазок |
Точка начала |
вилка |
|
|
сигналом для «вступления в игру» ДНК- |
|||||||||
с двумя вилками |
репликации |
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
геликазы. Сами ДНК-боксы не кодируют |
|||||
|
|
|
|
|
|
|
|
|
белок или РНК, хотя между ними |
|||||
|
|
|
|
|
|
|
|
|
располагаются отдельные гены. Продукты |
|||||
|
|
|
|
|
|
|
|
|
этих генов также большей частью вовлечены |
|||||
|
|
|
|
|
|
|
|
|
в «обслуживание» репликации ДНК. |
|||||
|
|
|
|
|
|
|
|
|
Инициация репликации не происходит, если |
|||||
Репликационная |
|
|
Репликационная |
|
||||||||||
вилка |
|
Точка начала |
вилка |
|
|
подавлен |
синтез белка. |
Порядок |
||||||
|
|
|
репликации |
|
|
|
||||||||
|
|
|
|
|
|
расположения |
ДНК-боксов, |
|||||||
Образование “репликационного глазка” с одной |
|
|||||||||||||
|
промежуточных областей и их количество |
|||||||||||||
или двумя репликационными вилками. Тонкие |
|
|||||||||||||
|
позволяет думать, что эволюционная |
|||||||||||||
стрелки указывают направление, в котором |
|
|||||||||||||
|
дивергенция OriC шла главным образом за |
|||||||||||||
расплетается родительская ДНК (из: Фаворова, |
|
|||||||||||||
|
|
|
|
|
|
|||||||||
1996, ñòð. 15). |
|
|
|
|
|
|
|
счетдупликацийитрипликаций.ОбластьOriC |
||||||
|
|
|
|
|
|
|
|
у E. сoli и B. subtilis, будучи лигированной с |
||||||
молекулы ДНК. Участок ДНК, в котором |
||||||||||||||
репликация уже завершилась, выглядит как |
фрагментами |
некоторых |
плазмид, |
|||||||||||
пузырек или “глазок” в нереплицированной |
превращается |
в «мини хромосому», |
||||||||||||
ДНК. Репликационные “глазки” образуются |
способную к автономной репликации (Из: |
|||||||||||||
в тех местах, где находятся точки начала |
Прозоров, 1998). |
|
|
|||||||||||
репликации (origin of replication). Они |
Для того, чтобы цепи ДНК |
|||||||||||||
разьединились, функционирует особый |
||||||||||||||
состоят примерно из 250-300 нуклеотидов. |
фермент - ДНК-геликаза, который |
|||||||||||||
Лучше изучены ориджины у E. coli и |
||||||||||||||
Bacillus subtilis. Область начала репликации |
связывается с белками, инициирующими |
|||||||||||||
хромосомы, OriC, включает в себя участки |
процессрепликации.Этотферментдвижется |
|||||||||||||
с так называемыми ДНК-боксами, и |
|
по одиночной цепи ДНК и, встречая участок |
||||||||||||
расположенными между ними короткими |
двойной спирали, разрывает водородные |
|||||||||||||
последовательностями. ДНК-боксы со |
связи между основаниями, разделяет цепи и |
|||||||||||||
специфическим “рисунком” нуклеотидов, |
продвигаетрепликационнуювилку. |
|
||||||||||||
преимущественно в 9 нп, перемежаются |
Субстратом для ДНК-полимеразы |
|||||||||||||
являютсядезоксирибонуклеозид-трифосфаты |
||||||||||||||
фрагментами в 12-13 п.н. с высоким |
|
|||||||||||||
содержанием АТ. Сами девятичленники |
(äÍÒÔ), |
полимеризующиеся |
íà |
|||||||||||
могут располагаться как в прямом, так и в |
одноцепочечной матрице. ДНК-полимеразы |
|||||||||||||
инвертированном порядке по отношению |
последовательнонаращиваютоднунитьДНК, |
|||||||||||||
друг к другу. Например, у B. subtilis в участке |
шаг за шагом присоединяя к ней следующие |
|||||||||||||
RимеетсяодиндевятичленникТТАТССАСА |
звенья в направлении от 5' к 3' концу, причем |
|||||||||||||
и два других девятичленных бокса, |
выборочередногодНТФдиктуетсяматрицей. |
|||||||||||||
ориентированных в противоположном |
|
Вклеткахприсутствуютнесколькоразныхтипов |
||||||||||||
112

Структура и организация генома |
Глава 6 |
|
|
ДНКполимераз, выполняющих различные функциииимеющихразноестроение:онимогут бытьпостроеныизразличногочислабелковых цепей (субъединиц), от одной до десятков. Однако, все они работают на любых последовательностях нуклеотидов матрицы; задачаэтихферментов-сделатьточнуюкопию каждойматрицы.
Генетическийматериалживыхорганизмов имеет огромные размеры и реплицируется с высокой точностью. В среднем, в процессе воспроизведения генома млекопитающего, состоящего из ДНК длиной 3 млрд. пар нуклеотидов,возникаетнеболеетрехошибок. При этом ДНК синтезируется чрезвычайно быстро(скоростьееполимеризацииколеблется в пределах от 500 нуклеотидов в секунду у бактерий до 50 нуклеотидов в секунду у млекопитающих).
Высокаяточностьрепликации,нарядусее высокойскоростью,обеспечиваетсяналичием специальных механизмов, осуществляющих коррекцию,т.е.устраняющихошибки.
Сутьмеханизмакоррекциизаключаетсяв том,чтоДНК-полимеразыдваждыпроверяют соответствие каждого нуклеотида матрице: один раз перед включением его в состав растущей цепи, второй раз перед тем, как включить следующий нуклеотид. Очередная фосфодиэфирная связь синтезируется лишь в томслучае,еслипоследнийнуклеотидрастущей цепи ДНК образовал правильную уотсонкриковскую пару с соответствующим нуклеотидомматрицы.
ДНК-полимеразы не могут начинать синтез ДНК на матрице, а способны только добавлять новые дезоксирибонуклеотидные звеньяк3'-концуужеимеющейсяполинуклео- тиднойцепи.Такуюзаранееобразованнуюцепь, ккоторойдобавляютсянуклеотиды,называют затравкой(илипраймером).Затравка состоит из РНК. КороткуюРНК-затравкусинтезирует из рибонуклеозидтрифосфатов фермент, не обладающийкорректирующейактивностьюи называемый ДНК-праймазой. Праймазная активность может принадлежать либо отдельному ферменту, либо одной из субьединицДНК-полимеразы.
Установлено, что дочерние цепи ДНК растуттольковнаправлении5'→ 3',т.е.всегда
удлиняется 3'-конец затравки, а матрица считываетсяДНК-полимеразойвнаправлении 3' → 5'. Синтез ДНК происходит непрерывно тольконаоднойизматричныхцепей.Навторой цепи ДНК синтезируется сравнительно короткими фрагментами - от 100 до 1000 нуклеотидов, названными “фрагментами Оказаки” по имени открывшего их ученого - ТунекоОказаки(Рис.6.10).
СодиночнымицепямиДНКсвязываются специальные белки, дестабилизирующие спираль (SSB - single strand binding proteins). Они не позволяют им сомкнуться. Для того, чтобы репликационная вилка могла продвигаться вперед, вся еще не удвоенная часть ДНК должна была бы очень быстро вращаться.БелкиДНК-топоизомеразывносят одноцепочечныеилидвуцепочечныеразрывы, позволяющиецепямДНКразделиться,азатем заделываютэтиразрывы.
На Рис. 6.11 показано расположение цепей и молекул ферментов во время репликации.
Спираль расплетается с помощью ДНК-геликазы; этому процессу помогают ДНК-топоизомераза, раскручивающая цепи ДНК, и множество молекул
Рисунок 6.10
|
|
3’ |
|
|
5’ |
|
|
Ведущая |
5’ |
|
öåïü |
|
3’ |
|
|
|
|
|
5’ |
РНК-затравки |
|
|
|
3’ |
|
|
|
|
Отстающая |
|
|
öåïü |
|
|
3’ |
|
|
5’ |
Строение репликационной вилки. Направление синтеза ДНК совпадает с направлением расплетания двойной спирали лишь для одной из новосинтезированных цепей - ведущей. Вторая цепь - отстающая - синтезируется прерывисто, в виде коротких фрагментов Оказаки. В результате обе дочерние цепи растут в направлении от 5' к 3' (из: Фаворова, 1996, стр. 15).
113

Глава 6 |
Структура и организация генома |
|
|
Рисунок 6.11
Матрица для синтеза ведущей цепи
ДНК-полимераза на ведущей цепи
Новосинтезированная |
Топоизомераза |
|
öåïü |
||
|
Синтез следующего фрагмента Оказаки начинается здесь
ДНК-полимераза,  заканчивающая синтез фрагмента Оказаки на
заканчивающая синтез фрагмента Оказаки на  ДНК-геликаза
ДНК-геликаза
отстающей цепи
Дестабилизирующий Праймаза белок
РНК-затравка
Матрица для синтеза отстающей цепи
Направление движения репликационной вилки
Расположение основных белков в репликационной вилке (из: Фаворова, 1996, стр. 16).
дестабилизирующего белка (SSB), |
цепями ДНК. В области вилки действуютдве |
||||
связывающихся с обеими одиночными |
ДНК-полимеразы - на ведущей и отстающей |
||||
|
цепи. На ведущей цепи ДНК-полимераза |
||||
Дополнение 6.6 |
работает непрерывно, а на отстающей |
||||
Все живые организмы на Земле обычно |
ферментвремяотвременипрерываетивновь |
||||
делят на прокариот и эукариот (от греч. карион |
возобновляет свою работу, используя |
||||
- ядро). Главной особенностью прокариот |
|||||
короткие РНК-затравки, синтезируемые |
|||||
является отсутствие у них в отличие от эукариот |
|||||
ДНК-праймазой. Молекула ДНК-праймазы |
|||||
(эу - по-гречески - истинный) полноценного |
|||||
непосредственно связана с ДНК-геликазой, |
|||||
клеточного ядра, покрытого оболочкой. |
|||||
образуя |
структуру, |
называемую |
|||
Генетический материал прокариот расположен |
|||||
праймосомой. Праймосома движется в |
|||||
в нуклеоиде - примитивном эквиваленте ядра |
|||||
направлении раскрывания репликационной |
|||||
эукариот. Клетки прокариот имеют очень |
|||||
|
|
|
|
||
небольшие размеры - около 1мкм. Объем |
вилки и по ходу движения синтезирует РНК- |
||||
эукариотических клеток в 800-1000 раз больше |
затравку для фрагментов Оказаки. В этом же |
||||
объема клеток прокариот. К прокариотам |
направлении движется ДНК-полимераза |
||||
относятся бактерии и археи (или архебактерии), |
ведущейцепииДНК-полимеразаотстающей |
||||
предки которых возникли около 4 млрд. лет |
цепи. Для этого, как полагают, последняя |
||||
назад. В последнее время все чаще говорят о |
накладывает цепь ДНК, которая служит ей |
||||
трех царствах живых существ: бактериях, |
|||||
матрицей, саму на себя, что и обеспечивает |
|||||
археях и эукариотах. Эукариотымогутбытькак |
|||||
разворот ДНК-полимеразы отстающей цепи |
|||||
одноклеточными так и многоклеточными. Они |
|||||
на 180 градусов (Рис. 6.12). Согласованное |
|||||
появились на Земле примерно через 500 млн. |
|||||
движение двух молекул ДНК-полимераз |
|||||
лет после прокариот (см. Кулаев, 1998; |
|||||
обеспечивает |
координированную |
||||
Алиханян и др., 1985, с.435,438). |
|||||
|
|
|
|
||
|
|
|
|
|
|
114

Структура и организация генома |
Глава 6 |
|
|
Рисунок 6.12
Репликация ДНК (“модель тромбона”) (Из: Russell, 1998, p. 357).
репликацию обеих нитей. В конце процесса ДНК – лигаза катализирует формирование фосфодиэфирной связи между 3’-OH и 5’– фосфатных групп во вновь синтезированной цепиДНК.
Всего в репликационной вилке одновременно работает около двадцати разных белков (на Рис. 6.11 и 6.12 показана только часть из них). Этот комплекс ферментов называют реплисомой.
Процесс репликации хромосомы бактерий начинается в точке начала репликации и продолжается до тех пор, пока не удвоится вся ДНК хромосомы.
6.3.2. Особенности репликации ДНК у эукариот
Молекулярно - биологиче ские процессы, происходящие во время репликации ДНК, в основном похожи у эукариот и прокариот. При этом обнаружены следующиеразличия:во-первых,репликация ДНКуэукариотпроисходитнаопределенной стадии клеточного цикла, во-вторых, если бактериальная хромосома представляет собой единицу репликации - репликон, то репликация ДНК эукариотической хромосомы осуществляется посредством
разделения ее на множество отдельных репликонов.
Клеточные циклы у эукариот качественно не различаются у разных видов и при делении клеток в разных тканях одного вида. Замечены различия, главным образом, в длине цикла. Среди высших эукариот некоторые клетки делятся через 10 минут, другие через 3 часа, третьи – через 200 часов.
Клеточный цикл у большинства соматических клеток высших эукариот подразделяют на 4 стадии: G1 (gap1, предсинтетический период подготовки к синтезу ДНК), S (Synthesis, период синтеза ДНК), G2 (gap 2, постсинтетический период подготовки к клеточному делению) и M (mitosis, собственно процесс клеточного деления). В культуре клеток человека весь цикл занимает примерно 24 часа, при этом на G1, S, G2 и M стадии приходится 10, 9, 4 и 1 час соответственно. G1, S è G2 фазы вместе составляют интерфазу. Наиболее детальные данные получены при изучении клеточных циклов дрожжей (Рис. 6.13). Данные генетических и молекулярных исследований показали,чтовклеточныхциклахсуществует ряд этапов, на которых осуществляется
115

Глава 6 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Структура и организация генома |
||
|
|
|
|
|
|||||||||||||
контрольпродвиженияклеткиотоднойфазы |
|
клетка не выросла до нормальных размеров, |
|||||||||||||||
к другой – стадии проверки (checkpoints). |
|
и окружающая среда недостаточно хороша, |
|||||||||||||||
Первая стадия проверки, называемая у |
|
клетка неспособна перейти к стадии M. |
|||||||||||||||
дрожжей START (у млекопитающих – G1 |
|
Третья проверка происходит в течение |
|||||||||||||||
checkpoints). Если клетка не выросла до |
|
фазы M: хромосомы должны быть надежно |
|||||||||||||||
необходимых размеров и окружающая среда |
|
прикреплены к нитям митотического |
|||||||||||||||
недостаточно хороша, |
|
клетка |
|
будет |
|
веретена, чтобы начать разделение хроматид. |
|||||||||||
оставаться в G1. |
|
|
|
|
|
|
|
|
|
|
|
|
|
Ключевыми |
компонентами, |
||
|
В S-фазе разные участки генома |
|
вовлеченными в регуляторные события в |
||||||||||||||
реплицируются, по-видимому, в разное |
|
чек-пойнтах, являются белки, известные под |
|||||||||||||||
время. В культуре клеток человека сначала |
|
названием циклинов (Cyclins) и циклин- |
|||||||||||||||
синтезируется ДНК, которая выявляется в R- |
|
зависимыхкиназ(Cdk’s).УдрожжейкаквG1, |
|||||||||||||||
бэндах метафазных хромосом (см. Раздел |
|
такивG2 –стадияхпроверкифункционирует |
|||||||||||||||
9.4.3.), которые обогащены генами. В конце |
|
одна и та же Cdk, у млекопитающих - две |
|||||||||||||||
S-периода синтезируется ДНК G-бэндов. |
|
разных. Специфичность каждой из стадий |
|||||||||||||||
Полагают, что между этими отрезками S- |
|
проверки определяется типом участвующих |
|||||||||||||||
периода существует стадия проверки. |
|
|
|
|
в этом циклинов. |
|
|||||||||||
|
Другая стадия проверки – G2 – |
|
 G1 один или более G1-циклинов |
||||||||||||||
находится на границе G2 è M. Åñëè íå |
|
связывается с Cdk (CDC28/cdc2 киназой) и |
|||||||||||||||
завершилась репликация всей ДНК, если |
|
активирует ее. Затем Cdk фосфорилирует |
|||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
ключевые белки, необходимые для |
|
|
Рисунок 6.13 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Образование комплекса G1 циклина и Cdk; |
|
|
|
|
|
|
перехода в S-период. Как только |
|||||||||
|
Cdk фосфорилирует белки, необходимые |
|
|
|
|
|
|
|
циклин активирует Cdk, уровень |
||||||||
|
для перехода клетки в S-период |
|
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
Деградация |
циклинов уменьшается из-за |
|||
|
циклин |
|
|
|
|
|
|
|
|
|
|
|
G1 |
циклина |
усиления процессов протеолиза. |
||
|
(CDC28/cdc2)-зависимая |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
киназа (Cdk) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Аналогичный процесс |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
G1 циклин |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
протекает на стадии проверки G2, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
когда один или болеемитотических |
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
II I I I I |
I I |
I |
I |
|
|
|
|
циклинов |
связывается с Cdk с |
|||
|
|
|
|
|
I |
|
|
|
образованием MPF (M-phase |
||||||||
|
|
|
|
|
I |
|
|
|
|
|
|
I |
|
|
|||
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
I |
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
promoting factor). Затем, когда |
|||
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
||||
|
|
|
I |
I |
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
I |
|
|||||
|
|
I |
|
|
|
|
|
|
|
|
|
I |
другие ферменты фосфорилируют |
||||
|
I |
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
I |
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
||||
|
I |
|
|
|
|
|
|
|
|
|
|
|
S |
|
I |
и дефосфорилируют его, MPF |
|
|
I |
|
|
|
G1 |
|
|
|
|
|
|
|
|
|
I |
||
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
активируется и стимулирует |
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
||
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
события, |
необходимые для |
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
I |
I |
перехода клетки в митотическую |
||
|
I |
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
M |
|
|
|
|
|
I I |
|
|||||||
|
|
I |
|
|
G2 |
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
стадию. |
|
|||||||
|
|
|
I |
|
|
|
|
|
|
|
|
||||||
|
|
|
|
I |
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
I |
|
|
В ходе митоза, сразу после |
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
I |
|
|
||
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
||||
|
|
|
|
|
I |
I |
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
I |
I I |
I |
I |
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
I I I I |
|
|
|
|
|
|
|
метафазы, митотический циклин |
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
деградирует, что приводит к |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
инактивации MPF, позволяющей |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
M-циклин |
клетке завершить митоз. |
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
Деградация M-циклинов |
|
|
|
|
|
|
|
|
|
|
|
У дрозофилы в ходе раннего |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
эмбрионального развития (первые |
|
|
Митотические циклины связываются с Cdk, |
два часа после оплодотворения |
|||||||||||||||
|
образуя MPF (M-phase promoting factor), который |
||||||||||||||||
|
яйцеклетки сперматозоидом) ядра |
||||||||||||||||
|
затем активируется и стимулирует цепь событий, |
||||||||||||||||
|
продвигающих клетку в M-фазу |
|
|
делятся каждые 9.6 мин. (при 240C). |
|||||||||||||
|
Некоторые события, контролирующие прохождение через |
||||||||||||||||
|
Так как интерфаза в этих делениях |
||||||||||||||||
|
клеточные циклы у дрожжей (Из: Russell, 1998, P. 362). |
|
|
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|