

Клеточная сигнализация
В многоклеточных организмах поддержание гомеостаза обеспечивают 3 системы:
1). нервная, 2). гуморальная, 3). иммунная.
Регуляторные системы функционируют с участием сигнальных молекул.
Сигнальные молекулы– это органические вещества, которые переносят информацию.
К сигнальным молекулам относятся гормоны, нейромедиаторы, факторы роста, цитокины и эйкозаноиды.
ЦНС для передачи сигнала использует нейромедиаторы, гуморальная система – гормоны, иммунная - цитокины.
Гормоны, это сигнальные молекулы беспроводного системного действия.
Отличием истинных гормоновот других сигнальных молекул, является то, что они синтезируются в специализированных эндокринных клетках, транспортируются кровью и действуют дистантно на ткани мишени.
Гормоны по строению делятся: на
белковые(гормоны гипоталамуса, гипофиза),
производные аминокислот(тиреоидные, катехоламины)
и стероидные(половые, кортикоиды).
Пептидные гормоны и катехоламины растворимы в воде, они регулируют преимущественно каталитическую активность ферментов.
Стероидные и тиреоидные гормоны водонерастворимы, они регулируют преимущественно количество ферментов.
Гормоны влияют на активность и количество ферментов в клетке не напрямую, а через каскадные системы(аденилатциклазную, гуанилатциклазную, инозитолтрифосфатную,RASи т.д.),состоящие из:
рецепторов;
регуляторных белков (G-белки,IRS, Shc, STAT и т.д.).
вторичных посредников, (messenger - посыльный) (Са2+, цАМФ, цГМФ, ДАГ, ИТФ);
ферментов (аденилатциклаза, фосфолипаза С, фосфодиэстераза, протеинкиназы А, С,G, фосфопротеинфосфотаза);
Необходимость каскадных систем связана с тем, что, во-первых, водорастворимые гормоны не проходят клеточную мембрану, во-вторых, эти системы обеспечивают усиление первичного сигнала гормонов в миллионы раз. В результате даже одна молекула гормона способна активировать миллионы ферментов и вызвать метаболический эффект.
Водонерастворимые гормоны самостоятельно проходят клеточные мембраны и реализуют свой эффект с участием цитоплазматических и ядерных рецепторов.
Рецепторы
Рецепторы- это белки, встроенные в клеточную мембрану или находящиеся внутри клетки, которые, взаимодействуя с сигнальными молекулами, меняют активность регуляторных белков.
По локализации рецепторы делятся на: 1) цитоплазматические; 2) ядерные; 3) мембранные.
По эффекту рецепторы делятся на:активаторные(активируют каскадные системы) иингибиторные(блокируют каскадные системы).
Участие рецепторов в трансмембранной передаче сигнала
вторичные
посредники:
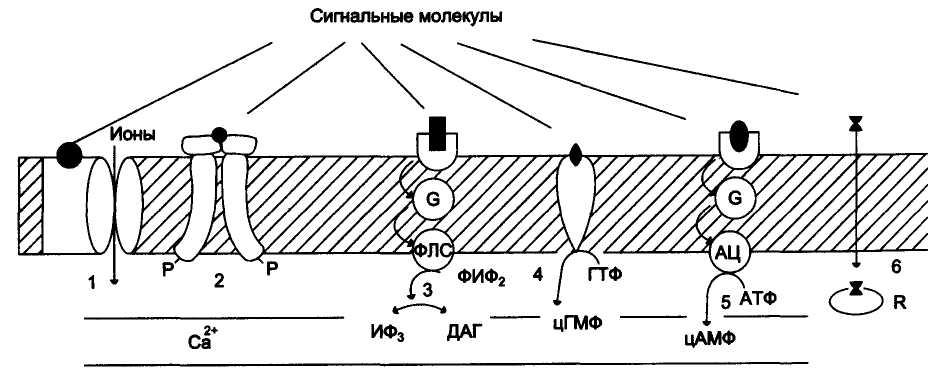
2). Рецепторы, с тирозинкиназной активностью (рецептор инсулина);
3). Рецепторы, активирующие инозитолтрифосфатную систему (α1-адренорецептор - у гепатоцитов);
4). Рецепторы, с гуанилатциклазной активностью (гуанилатциклаза, рецептор ПНФ);
5). Рецепторы, активирующие аденилатциклазную систему (β-адренорецепторы);
6). Рецепторы, связывающие гормон в цитозоле или ядре (рецептор кортизола).
По механизму передачи сигнала рецепторы делятся на 4 типа:
1). Рецепторы, связанные с ионными каналами. Это интегральные мембранные белки, состоящие из нескольких субъединиц, полипептидная цепь которых несколько раз пересекает клеточную мембрану. Они имеют центр связывания сигнальной молекулы и ионный канал. При связывании с сигнальными молекулами у этих рецепторов открываются или закрываются ионные каналы. Действуют они очень быстро в течение миллисекунд.
Эти рецепторы обеспечиваю синаптическую передачу в электрически возбудимых клетках. Например, катионные ацетилхолиновые никотиновые рецепторы скелетных мышц.
2). Рецепторы, с ферментативной активностью. Имеют разнообразное строение, регулируют клеточное деление, дифференцировку, развитие иммунного ответа.
Рецепторы с ферментативной активностью бывают 3 видов:
а). Рецепторы, с тирозинкиназной активностью(тирозиновые протеинкиназы). Это каталитические рецепторы, фосфорилирующие по тирозину белки-мишени. Их активируют инсулин, макрофагный колониестимулирующий фактор, тромбоцитарный производный фактор роста.
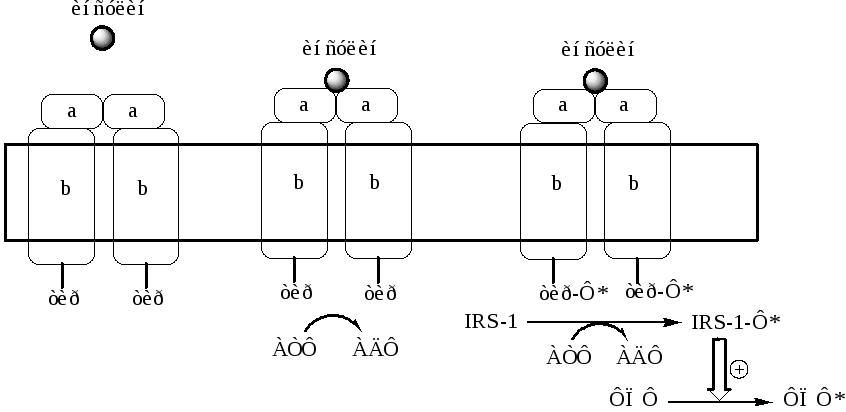
Например, мембранный рецептор инсулина, он является гликопротеином, который состоит из 2 α и 2 β субъединиц связанных дисульфидными связями. α субъединицы связывают инсулин, а β субъединицы обладают тирозинкиназной активностью. После присоединения гормона к α субъединицам, β субъединицы сначала фосфорилируют друг друга, а затем белок IRS-1 (insulin receptor substrate), который активирует функциональные ферменты (фосфопротеинфосфатаза). ФПФ в свою очередь дефосфорилирует и инактивирует инсулиновый рецептор.
б). Рецепторы, с фосфатазной активностью(тирозиновые протеинфосфотазы). Это каталитические рецепторы, дефосфорилирующие по тирозину белки-мишени (например, ФПФ).
в). Рецепторы с гуанилатциклазной активностью (гуанилатциклазы ГЦ). Это каталитические рецепторы, превращающие ГТФ в цГМФ, есть в сердце, легких, почках, надпочечниках, эндотелии кишечника, сетчатке и т.д.
Эти рецепторы находятся на мембране и в цитоплазме:
Мембранная ГЦ – гликопротеин (180кДа), имеет 3 домена: внеклеточный рецепорный, трансмембранный и внутриклеточный каталитический. Активируетсяпредсердным натрийуретическим фактором(АНФ), термостабильным токсином грамотрицательных бактерий, эндотелийпроизводным фактором, ацетилхолином+Са2+, серотонином, гистамином и т.д. Существует 3 вида.
Цитоплазматическая ГЦсостоит из α и β субъединиц и содержит гем, активируется оксидом азотаNO(а также Н2О2, О2, жирными кислотами и продуктами ПОЛ).
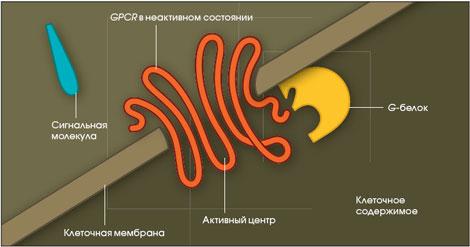
3). Рецепторы, сопряженные с G-белками (GPCRот англ. G – protein coupled receptor), по строению их еще называютсерпантинными.
|
Это мономерные интегральные мембранные белки, полипептидная цепь которых 7 раз пронизывает клеточную мембрану. Внеклеточный домен GPCRобеспечивает взаимодействие с гормоном, а внутриклеточный - контакт сG-белками. В настоящее время открыто более 200 видов GPCR. К GPCRотносятся: α иβрецепторы. К GPCRприсоединяются: адреналин (рецепторы α1и α2, β1и β2), ацетилхолин (рецепторы М1, М2, М3, М4), серотонин (1А, 1В, 1С, 2), дофамин (Д1и Д2), АКТГ, ТТГ, ФСГ, ЛГ, хорионический гонадотропин, простагландины, гастрин, холецистокинин, нейропептид Y, нейромедин К, вазопрессин, ангиотензин, вещество К, вещество Р, или нейрокинин 1, 2 и 3 типа, тромбин, интерлейкин-8, глюкагон, кальцитонин, секретин, соматолиберин, ВИП, гипофизарный аденилатциклазактивирующий пептид, глютамат (MG1 – MG7), аденин. |
|
|
|
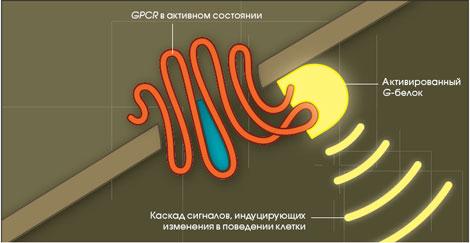
GPCR через G-белки активируют аденилатциклазную и инозитолтрифосфатную системы. GPCR инактивируются:
путем ухода сигнальной молекулы из активного центра рецептора;
путем фосфорилирования рецепторов протеинкиназами и последующего присоединения специального белка (например, β-аррестин);
GPCR участвуют почти во всех жизненно важных процессах, протекающих в организме: они поддерживают работу сердца, органов пищеварения и дыхания, мозговую деятельность.
Примерно половина современных лекарств взаимодействуют с GPCR.
4). Ядерные и цитоплазматические рецепторы. Находятся в цитоплазме или ядре, при взаимодействии с гормоном, образуют комплекс, который присоединяется к регуляторной нуклеотидной последовательности в ДНК, изменяет доступность промотора для РНК-полимеразы, соответственно меняет скорость транскрипции мРНК и трансляции новых ферментов.
Ядерные и цитоплазматические рецепторы содержат ДНК-связывающий домен, характеризующийся наличием двух структур «цинковых пальцев». Особенностью цитоплазматических рецепторов является связь с белком-шапероном, который стабилизирует их структуру.
Разные клетки организма, в зависимости от функций, имеют определенный набор рецепторов. На мембране одной клетки может быть более десятка разных типов рецепторов.