

Опыты по прививкам с яйцами морских ежей
Различные комбинации половинок двух яиц морского ежа Paracentrotus могут быть получены следующим образом. После удаления яиц из их оболочки им дают развиваться до 16-клеточной стадии. Наличие микромеров на вегетативном полюсе дает затем возможность определить плоскость разделения групп бластомеров. Для того чтобы сделать это, яйца за десять минут до разделения, которое производится с помощью стеклянной иглы, помещаются в морскую воду, не содержащую кальция.
|
|
|
Рис. 117. Схема, показывающая, как получается сращивание вместе бластомеров морского ежа (по Герстадиусу). |

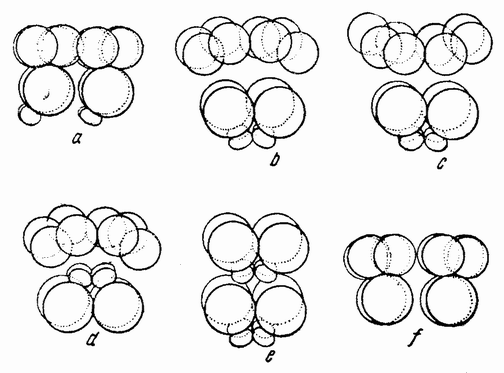
Когда полярность обеих соединяемых половинок одинакова, как показано на рис. 118 a, из них развивается нормальный плутеус. Так как разделение могло пройти либо через проспективную среднюю плоскость, если таковая в это время уже определена, либо под прямым углом к этой средней плоскости, то, повидимому, в это время такая плоскость отсутствует или, если присутствует, то может быть изменена и установлена заново. Когда половинки соединяются так, как показано на рис. 118b, т. е. с обратной полярностью, то вегетативная область каждой половинки инвагинируется, как изображено на рис. 119аиb. В этом случае образуется плутеус с двумя совершенно разделенными пищеварительными трактами, с двумя лежащими друг против друга анальными отверстиями, двумя средними и передними кишками и с одним общим ротовым отверстием (рис. 119с). Скелет почти удвоен. {203}
Если половинки соединены, как изображено на рис. 118 с, т. е. ориентированы под прямым углом друг к другу, то гаструляция начинается в двух точках и два первичных кишечника раньше или позже соединяются, образуя раздвоенную или единую кишку. В этом случае развивается либо типичный, либо, более часто, ненормальный плутеус.
|
|
|
Рис. 118. Схема различных способов соединения половинок 16-клеточных стадий морского ежа (по Герстадиусу). |

При соединении анимальных половинок, показанном на рис. 118 d, развивается бластула с двумя рядами ресничек, идущими в противоположных направлениях. Если соединены (рис. 118е) меридиональная и анимальная половинки, то комбинация состоит из 12 мезомеров (эктодерма), 2 макромеров и 2 микромеров и развивается типичный плутеус (рис. 120с). {204}
Если анимальная половинка присоединена к анимальному же концу целой 16-клеточной стадии (рис. 118 f), то такая комбинация содержит две трети эктодермы и одну
|
|
|
Рис. 119. а— гаструляция соединенных половинок (рис. 118);b— то же самое у другого подобного же соединения;с— развившийся двойной плутеус (по Герстадиусу). |
|
|
|
Рис. 120. а— атипичный плутеус, образовавшийся в результате соединения, изображенного на рис. 121с;b— плутеус, развившийся из соединения изображенных на рис. 121d;с— плутеус из комбинацииерис. 121 (по Герстадиусу). |


треть энтодермы, из нее может развиться плутеус, размеры которого будут крупнее нормального.
В случае соединения вегетативной половинки с анимальным концом целой 16-клеточной стадии (как показано на рис. 118 d) получается один плутеус с двумя кишечниками и одним ртом, сходный с изображенным на рис. 119с. {205}
При соединении двух вегетативных половинок (рис. 118 h) происходит две инвагинации, причем личинки оказываются при этом совершенно ненормальными.
Четыре микромера имплантировались в другую 16-клеточную стадию (рис. 118 i), образованную из двух половинок, как изображено на рис. 118а. Число мезенхимных клеток на стадии гасгрулы было удвоенным. Развивались типичные личинки без удвоения скелетных элементов.
В предыдущих случаях внутренние поверхности соединенных половинок были направлены друг к другу.
|
|
|
Рис. 121. Схема различных способов соединения половинок 16-клеточных стадий морских ежей (по Герстадиусу). |
В другой серии опытов наружная поверхность одной половинки обращалась к внутренней поверхности другой половинки (рис. 121 а). Для того чтобы понять, как эта система выправляется, полезны будут следующие схемы. На рис. 122анарисована нормальная бластула, у которой вегетативный полюс заштрихован. Если эту половинку отрезать и повернуть так, чтобы ее поверхность расположилась напротив внутренней поверхности верхней половинки (рис. 122b), то обе половинки изгибаются, как изображено на рис. 122с. Затем две полости соединяются путем удаления перегородки (122d) и получается единая полость (е—f). Внешние поверхности обеих {206} половинок находятся теперь на внешней поверхности одной бластулы, но здесь имеется два центра вегетативных клеток, лежащих на экваторе друг против друга.
|
|
|
Рис. 122. Схемы, показывающие результат соединения вегетативных половинок, повернутых и соединенных с анимальной половинкой так, как показано на рис. b(по Герстадиусу). |
|
|
|
Рис. 123. Схема, показывающая результат соединения меридиональных половинок 16-клеточных стадий, как показано на рис. b(по Герстадиусу). |


На рис. 123 апредставлены две разделенные по анимальной оси половинки, одна из которых перевернута и {207} ее внешняя поверхность помещена внутрь другой половинки (123b). Обе половинки закругляются, как показано на рис. 123с. Перегородка удаляется (d) и получается единая полая бластула. Вегетативная область находится на той же стороне бластулы, но несколько в сторону от нее (d—f).
С помощью этих схем легко понять поведение соединенных половинок 16-клеточной стадии. Если две половинки соединены, как на рис. 121 а, получаются как типичные, так и атипичные плутеусы. Если две половинки соединены, как на рис. 121с, то развиваются тоже и типичные и атипичные плутеусы. При обратной комбинации (рис. 121d) развитие идет совершенно неправильно (рис. 120b). Некоторая регуляция в этих случаях происходит, но гаструляция нарушается и плутеусы оказываются ненормальными в отношении энтодермы (рис. 120b).
Если восемь клеток двух вегетативных полушарий соединяются с той же ориентировкой (рис. 121 b), то происходит две инвагинации, а получающиеся плутеусы оказываются очень ненормальными.
Если с каждой из двух половинок удаляются микромеры и половинки соединяются затем так, как изображено на рис. 121 f, то не образуется первичной мезенхимы, а с опозданием появляется вторичная мезенхима. В этом случае может развиться нормальный плутеус.
Эти опыты прививок показывают, что верхняя и нижняя половинки уже специфичны в отношении их будущего развития. Этого можно было ожидать из поведения анимальной и вегетативной половинок при изолировании их, а также и из поведения анимальных и вегетативных половинок несегментированных яиц. Существует некоторое взаимное восстановление клеток, но в целом при наличии двух групп макромеров очевидна тенденция удвоения (что дает начало первичному кишечнику). В какой степени макромеры действуют в качестве индуктора для остального эмбриона, не вполне ясно. Факты говорят скорее противоположное.
|
<< |
|
{208} |
|
>> |