

5.4. Обмен и роль кетоновых тел
Синтез кетоновых тел происходит только в митохондриях печени (рис.9). Источником для синтеза кетоновых тел служит ацетил-КоА, образующийся при окислении жирных кислот. Таким образом, кетоновые тела – это вещества липидного происхождения, но в отличие от липидов, они являются веществами гидрофильной природы, поэтому в плазме транспортируются в свободном виде. При синтезе кетоновых тел из 3 молекул ацетил-КоА образуется -гидрокси--мелилглутарил-КоА (ГМГ-КоА). Далее ГМГ-КоА-лиаза расщепляет ГМГ-КоА на ацетоацетат (-кетобутират) – первое кетоновое тело и ацетил-КоА. -кетобутират может восстанавливаться при участии НАДН до -гидроксибутирата (второе кетоновое тело, преобладает в крови). При высокой концентрации ацетоацетата часть его неферментативно декарбоксилируется, превращаясь в ацетон. Ацетон не утилизируется тканями, выделяется с выдыхаемым воздухом, мочой, потом. Таким путем организм удаляет избыточное количество кетоновых тел, которые (кроме ацетона), являясь водорастворимыми кислотами, вызывают ацидоз. Кетоновые тела являются дополнительными источниками энергии при голоде и стрессе, особенно для головного мозга, который не использует жирные кислоты как источник энергии. Также кетоновые тела используют скелетные мышцы, сердце, почки. При этом -кетобутират превращается в две молекулы ацетил-КоА, сгорающие в ЦТК и ДЦ (при этом каждая молекула ацетил-КоА дает 10 АТФ). Кроме того, кетоновые тела могут использоваться как субстраты для синтеза липидов головного мозга и периферических нервов.
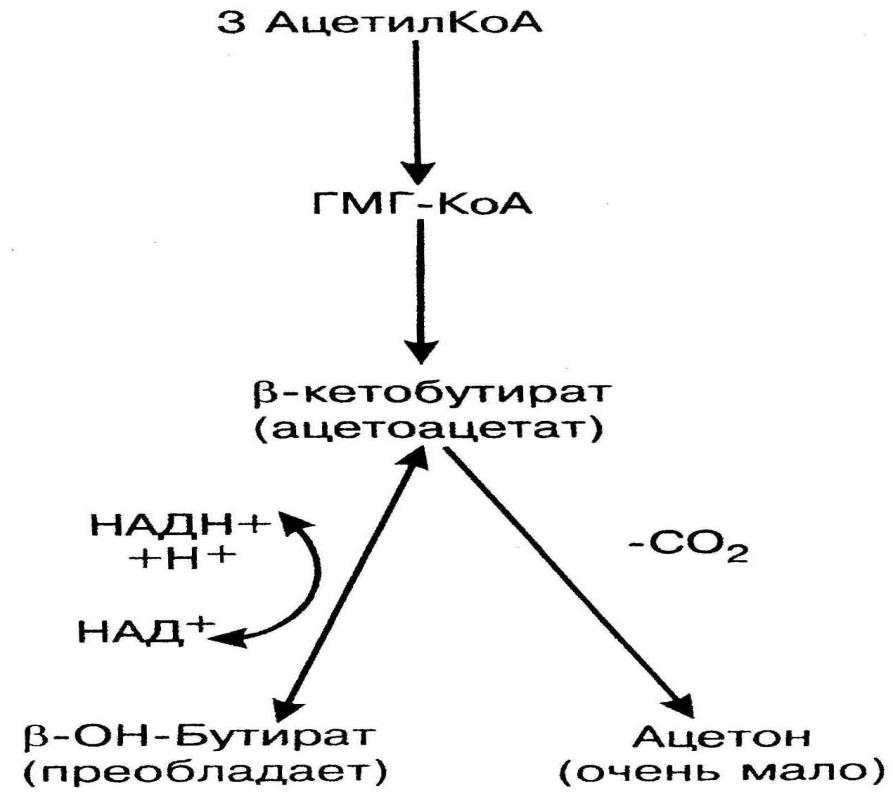
Рис.9. Кетогенез
В норме концентрация кетоновых тел в крови низкая (0,1 – 0,3 ммоль/л), но при ряде ситуаций она увеличивается, это называют гиперкетонемией, при этом избыток кетоновых тел выделяется с мочой – кетонурия. Все вместе называют термином кетоз. Кетозы бывают физиологическими (при голодании, тяжелой работе), патологическими (при лихорадках, неукротимой рвоте). Особой формой кетоза является диабетическая кома (концентрация кетоновых тел в крови может повышаться до 12-35 ммоль/л), при этом развивается кетоацидоз.
5.5. Обмен, роль и транспорт холестерина
В сутки в организме синтезируется около 1 г холестерина (рис.10). Основное место синтеза – печень (до80%), меньше синтезируется в кишечнике, коже и других тканях. С пищей поступает около 0,4 г холестерина, его источником является только пища животного происхождения. Холестерин необходим для построения всех мембран, в печени из него синтезируются желчные кислоты, в эндокринных железах – стероидные гормоны, в коже – витамин Д.
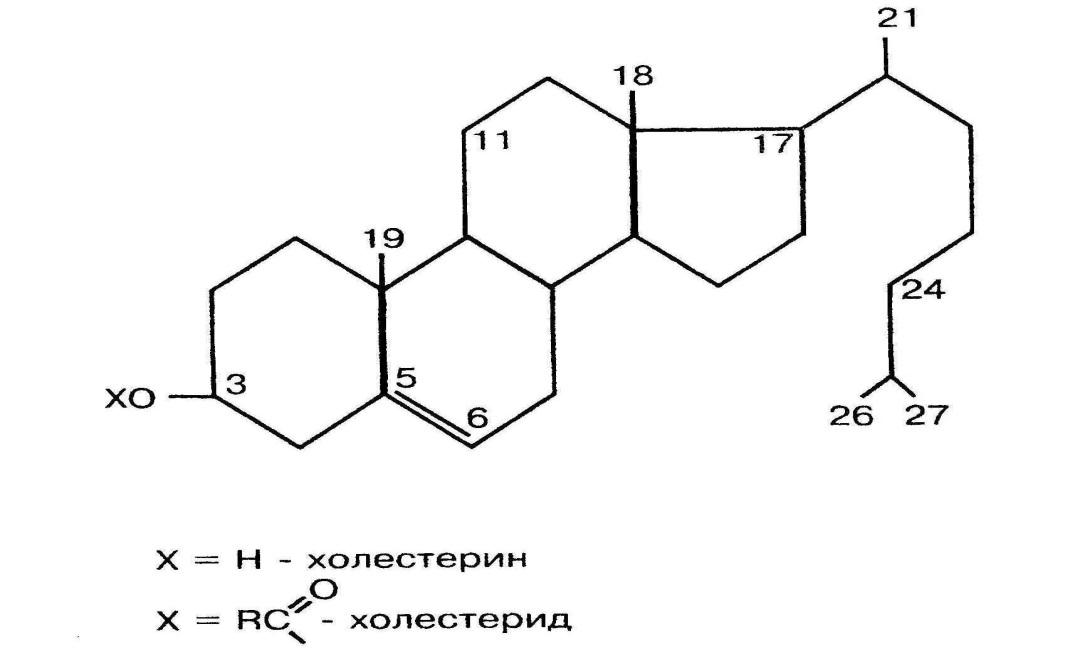
Рис.10 .Холестерин
Сложный путь синтеза холестерина можно поделить на 3 этапа (рис.11). Первый этап заканчивается образованием мевалоновой кислоты. Источником для синтеза холестерина служит ацетил-КоА. Сначала из 3 молекул ацетил-КоА образуется ГМГ-КоА – общий предшественник в синтезе холестерина и кетоновых тел (однако реакции синтеза кетоновых тел происходят в митохондриях печени, а реакции синтеза холестерина – в цитозоле клеток). Затем ГМГ-КоА под действием ГМГ-КоА-редуктазы восстанавливается до мевалоновой кислоты с использованием 2 молекул НАДФН. Эта реакция является регуляторной в синтезе холестерина. Синтез холестерина тормозит сам холестерин, желчные кислоты и гормон голода глюкагон. Усиливается синтез холестерина при стрессе катехоламинами.
На втором этапе синтеза из 6 молекул мевалоновой кислоты образуется углеводород сквален, имеющий линейную структуру и состоящий из 30 атомов углерода.
На третьем этапе синтеза происходит циклизация углеводородной цепи и отщепление 3 атомов углерода, поэтому холестерин содержит 27 углеродных атомов. Холестерин является гидрофобной молекулой, поэтому транспортируется кровью только в составе разных липопротеинов.

Рис. 11 Синтез холестерина
Липопротеины– липид-белковые комплексы, предназначенные для транспорта нерастворимых в водных средах липидов по крови (рис.12). Снаружи липопротеины (ЛП) имеют гидрофильную оболочку, которая состоит из молекул белков и гидрофильных групп фосфолипидов. водонерастворимых. ЛП делятся (по плотности и подвижности в электрическом поле) на 4 класса. Плотность ЛП определяется соотношением белков и липидов. Чем больше белка, тем больше плотность и тем меньше размер.

Рис.12. Строение липопротеидов
1 класс – хиломикроны (ХМ). Содержат 2%белка и 98% липидов, среди липидов преобладают экзогенные жиры, переносят экзогенные жиры от кишечника к органам и тканям, синтезируются в кишечнике, в крови присутствуют непостоянно – только после переваривания и всасывания жирной пищи.
2 класс – ЛП очень низкой плотности (ЛПОНП) или пре--ЛП. Белка в них 10%, липидов – 90%, среди липидов преобладают эндогенные жиры, транспортируют эндогенные жиры из печени в жировую ткань. Основное место синтеза – печень, небольшой вклад вносит тонкий кишечник.
3 класс – ЛП низкой плотности (ЛПНП) или -ЛП. Белка в них 22% , липидов – 78%, среди липидов преобладает холестерин. Нагружают клетки холестерином, поэтому их называют атерогенными, т.е. способствующими развитию атеросклероза (АС). Образуются непосредственно в плазме крови из ЛПОНП под действием фермента ЛП-липазы.
4 класс ЛП высокой плотности (ЛПВП) или -ЛП. Белка и липидов содержат по 50%, среди липидов преобладают фосфолипиды и холестерин. Разгружают клетки от избытка холестерина, поэтому являются антиатерогенными, т.е. препятствующими развитию АС. Основное место их синтеза – печень, небольшой вклад вносит тонкий кишечник.
Транспорт холестерина липопротеинами.Печень является основнымместом синтеза холестерина. Холестерин, синтезированный в печени, упаковывается в ЛПОНП и в их составе секретируется в кровь. В крови на них действует ЛП-липаза, под влиянием которой ЛПОНП переходят в ЛПНП. Таким образом, ЛПНП становятся основной транспортной формой холестерина, в которой он доставляется к тканям. ЛПНП могут попадать в клетки двумя путями: рецепторным и нерецепторным захватом. Большинство клеток на своей поверхности имеют рецепторы к ЛПНП. Образовавшийся комплекс рецептор-ЛПНП эндоцитозом попадает внутрь клетки, где распадается на рецептор и ЛПНП. Из ЛПНП при участии лизосомальных ферментов освобождается холестерин. Этот холестерин используется для обновления мембран, тормозит синтез холестерина данной клеткой, а также, если количество холестерина, поступающего в клетку, превышает ее потребность, то подавляется и синтез рецепторов к ЛПНП. Это уменьшает поток холестерина из крови в клетки, таким образом, клетки, для которых характерен рецепторный захват ЛПНП, имеют механизм, который ограждает их от избытка холестерина. Для гладкомышечных клеток сосудов и макрофагов характерен нерецепторный захват ЛПНП из крови. В эти клетки ЛПНП, а значит, и холестерин попадают диффузно, то есть, чем их больше в крови, тем больше их попадает в эти клетки. Эти разновидности клеток не имеют механизма, который ограждал бы их от избытка холестерина. В «обратном транспорте холестерина» от клеток участвуют ЛПВП. Они забирают избыток холестерина из клеток и возвращают его обратно в печень. Холестерин выводится с калом в виде желчных кислот, часть холестерина в составе желчи попадает в кишечник и также выводится с калом.