

3.2. Дыхательная цепь
В живых организмах окисление органических веществ кислородом происходит с образованием воды и СО2, что называется тканевым дыханием. Тканевое дыхание включает дегидрирование субстратов (отнятие от них водорода) и последующий многоэтапный процесс переноса электронов на кислород. Именно в ходе этого процесса происходит освобождение энергии, заключенной в электронах водорода. Однако в эксперименте непосредственное взаимодействие водорода с кислородом хотя и приводит к образованию воды, но сопровождается взрывом, ибо при этом вся энергия электронов водорода освобождается одномоментно. В организме образование воды происходит через ряд реакций, благодаря этому энергия электронов освобождается не одномоментно, а порциями. Это становится возможным благодаря тому, что во внутреннюю мембрану митохондрий встроена система переносчиков, называемая дыхательной цепью (ДЦ).
ДЦ – это совокупность дыхательных переносчиков, которые переносят сначала атомы водорода, а затем только электроны от субстрата на кислород. Большинство переносчиков представляют собой сложные белки-ферменты класса оксидоредуктаз. В их составе содержатся различные небелковые компоненты: ФМН, ионы Fе. Все компоненты ДЦ располагаются в порядке возрастания редокс-потенциала. Самый высокий редокс-потенциал у кислорода. Это обеспечивает последовательное перемещение электронов от НАДН на кислород, при котором на каждом этапе происходит освобождение энергии (рис. 2).
НАД2Н
ФМН2Н
Q2е
в2е
с12е
с2е
а2е
О









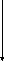

АТФ АТФ АТФ
2H+
H2О
О
2- 

Рис.2. Дыхательная цепь
Дегидрогеназы, отщепляющие атомы водорода от субстратов, делятся на НАД-зависимые и ФАД-зависимые. При участии НАД+-зависимых дегидрогеназ окисляются пируват, изоцитрат, α-кетоглутарат, малат, лактат и др. При участии ФАД-зависимых дегидрогеназ окисляются сукцинат, ацил-КоА, α-глицерофосфат.
Работа ДЦ по транспорту электронов на кислород представляет собой совокупность окислительно-восстановительных реакций, в которых каждый переносчик способен как окисляться, так и восстанавливаться. Так НАД+ присоединяет водород от субстратов и превращается в восстановленную форму – НАДН, который отдает атомы водорода на следующий переносчик – ФМН. При этом НАДН окисляется, а ФМН восстанавливается.
НАДН + Н+ + ФМН НАД+ + ФМНН2 и т. д.
Атомы Н, отщепляемые при участии ФАД-зависимых дегидрогеназ, поступают в ДЦ на уровне коэнзима Q (убихинона). До КоQ по дыхательной цепи одновременно передаются протоны и электроны водорода. Далее по системе цитохромов передаются только электроны. Цитохромы содержат гем, железо в котором может изменять свою валентность:
е +Fе3+Fе2+; Fe2+ - e Fe3+
Цитохром а, или цитохромоксидаза, передает электроны непосредственно на кислород, при этом цитохром а окисляется, а кислород, приняв 2 электрона, становится дважды отрицательно заряженным (О2-). Такой кислород взаимодействует с двумя протонами, отщепившимися на уровне КoQ, что приводит к образованию эндогенной воды. Но ДЦ работает не ради эндогенной воды, она работает ради освобождения энергии. В ДЦ освобождается энергия электронов водорода при переходе их с одного переносчика на другой. Около 60% энергии освобождается в виде тепла, а на тех участках, где значительный перепад редокс-потенциала и величина освобождаемой энергии составляет не менее 7,3 ккал, ее оказывается достаточно для синтеза АТФ. В виде АТФ в ДЦ аккумулируется 40% энергии. Участки, где в ДЦ происходит синтез АТФ, называются участками сопряжения. При этом происходит сопряжение (взаимосвязь) между окислением (т.е. переносом электронов по цепи переносчиков на кислород) и фосфорилированием АДФ до АТФ. Таких участков в ДЦ три:
Между ФМН и КоQ.
Между цитохромом в и с1.
Между цитохромом а и кислородом.
Если субстрат окисляется ФАД-зависимыми дегидрогеназами и атомы водорода вводятся в ДЦ на уровне КоQ, то в таком случае в ДЦ имеется два участка сопряжения: между цитохромом в и с1 и между цитохромом а и О. Результатом сопряжения в ДЦ является окислительное фосфорилирование (ОФ). ОФ – это синтез АТФ из АДФ и неорганического фосфата (Н3РО4) за счет энергии, освобождаемой при работе ДЦ. Количественно работу ДЦ оценивают по коэффициенту окислительного фосфорилирования Р/О. Коэффициент Р/О – это отношение числа молекул АТФ, образовавшихся в ДЦ, в расчете на один атом поглощенного кислорода. Если субстрат окисляется НАД+-зависимой дегидрогеназой, то Р/О равен 2,5, если ФАД-зависимой – то 1,5.
АТФ в клетке образуется из АДФ и неорганического фосфата. Для присоединения фосфата необходима энергия:
АДФ + Н3РО4 АТФ
В зависимости от того, какая энергия используется для синтеза АТФ, различают субстратное (СФ) и окислительное фосфорилирование (ОФ). В первом случае для синтеза АТФ используется энергия, возникшая при окислении субстрата, для него не нужна мембрана и при этом образуется мало АТФ. СФ протекает в два этапа:
Образование первичного макроэрга (сукцинил-КоА в цикле Кребса, 1,3-бисфосфоглицерата и фосфоенолпирувата в гликолизе)
Образование вторичного макроэрга – АТФ.
ОФ происходит только в митохондриях и для него необходима мембрана, при этом образуется большое количество АТФ. Для синтеза АТФ в ДЦ используется энергия электрохимического потенциала водорода (∆µН+), который возникает за счет выброса протонов из внутренней мембраны митохондрий в межмембранное пространство. Обратный процесс переноса протонов в матрикс через протонные каналы сопровождается освобождением энергии. Эту энергию и использует АТФ-синтаза для синтеза АТФ.
Полное окисление ацетил-КоА до СО2 и воды (т.е. в цикле Кребса и ДЦ) дает 10 АТФ, а полное окисление пирувата – 12,5 АТФ ( в окислении пирувата участвуют 4 НАД+-зависимых дегидрогеназы, что дает 4х2.5=10 АТФ, 1 ФАД-зависимая дегидрогеназа, что дает 1,5 АТФ и 1 АТФ образуется субстратным фосфорилированием в цикле Кребса. В окислении ацетил-КоА принимает участие на одну НАД+-зависимую дегидрогеназу меньше). Регуляция ДЦ. Скорость использования АТФ регулирует скорость потока электронов по ДЦ. Если АТФ не используется и его концентрация в клетке возрастает, то прекращается поток электронов к кислороду. И наоборот, расход АТФ и накопление АДФ увеличивает окисление субстратов и потребление кислорода. Зависимость интенсивности дыхания митохондрий от концентрации АДФ называют дыхательным контролем. Кроме того, работу ДЦ усиливают катехоламины, глюкагон и йодтиронины.
Разобщение в ДЦ. Некоторые химические вещества могут переносить протоны из межмембранного пространства через мембрану в матрикс, минуя протонные каналы. В результате этого снижается электрохимический потенциал и снижается синтез АТФ. Это явление называют разобщением окисления и фосфорилирования. В результате разобщения количество АТФ снижается, в результате чего будут страдать все процессы, требующие энергии АТФ (клинически проявляется слабостью, снижением работоспособности, повышением теплопродукции).В этом случае окисление НАДН и ФАДН2 возрастает, возрастает и количество поглощенного кислорода, но большая часть энергии выделяется в виде тепла (может повышаться температура тела) и коэффициент Р/О снижается : становится меньше 2,5 или 1,5 для НАД+- и ФАД-зависимых дегидрогеназ соответственно. Вещества, вызывающие разобщение, называются разобщителями. Физиологическими разобщителями являются жирные кислоты в буром жире. Кроме того, разобщение вызывают большие дозы йодтиронинов, токсические вещества, например, 2,4-динитрофенол; а также некоторые бактериальные токсины, например, дифтерийный. Разобщение окисления и фосфорилирования может быть биологически полезным. Оно позволяет генерировать тепло для поддержания постоянной температуры тела у новорожденных, у зимнеспящих животных и в процессе адаптации к холоду.