

Биология / Конспект лекции № 2
.docxКонспект лекции № 2.
Тема. Молекулярный уровень организации живого (нуклеиновые кислоты, липиды, углеводы) и энергетика клетки.
Нуклеиновые кислоты - это биологические гетерополимеры нуклеотидов.
Они имеют сложную первичную (цепочка нуклеотидов), вторичную (первичная спираль) и третичную структуру (вторичную спираль, кольцо, глобулу).
Первичная структура нуклеиновых кислот представляет собой цепочку из нуклеотидов.
Выделяют 8 разновидностей нуклеотидов. Четыре разновидности нуклеотидов для дезоксирибонуклеиновой кислоты (ДНК)-и четыре для рибонуклеиновой кислоты (РНК).
Каждый нуклеотид этих гетерополимеров состоит из 3 компонентов: азотистого основания, пятиуглеродного сахара (пентозы) и остатка фосфорной кислоты.
1.
Фосфорная кислота (Ф) одинакова у ДНК и
РНК.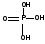
2. Пентоза - это пятиуглеродный моносахарид.
Пентоза
РНК - рибоза. Пентоза ДНК- дезоксирибоза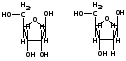
3. Азотистых оснований нуклеиновых кислот пять видов. Их разделяют на большие- производные пурина (два гетероциклических кольца) и маленькие- производные пиримидина (один гетероцикл).
пурин пиримидин
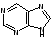
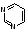
Пуриновых азотистых оснований два - аденин и гуанин (А и Г).
аденин гуанин
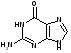
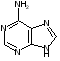
Пиримидиновых азотистых оснований три- тимин, цитозин и урацил (Т, Ц и У).
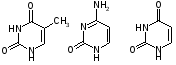
тимин цитозин урацил
Название нуклеотида определяется его азотистым основанием и часто сокращается до одной буквы.
В молекуле ДНК 4 вида нуклеотидов: А, Г, Т, Ц.
Аденин (аденин + фосфорная кислота + рибоза или АФД) = А. Гуанин (ГФД)- Г. Тимин (ТФД) = У. Цитозин (ЦФД) = Ц.
В молекуле РНК тоже 4 вида нуклеотидов. А, Г, У, Ц.
Аденин (аденин + фосфорная кислота + рибоза или АФР) = А. Гуанин (ГФР)- Г. Урацил (ТФР) = У. Цитозин (ЦФР) = Ц.
Пример
нуклеотида
РНК
-
АФР. Пример
нуклеотида ДНК - АФД
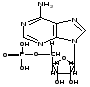
Таким образом, на этом уровне ДНК и РНК отличаются по виду остатка сахара (рибоза и дезоксирибоза) и по одному виду азотистого основания. В ДНК это тимин, в РНК урацил.
Молекулы нуклеиновых кислот представляют собой цепочки, соединенные посредством прочных ковалентных связей между фосфорной кислотой одного нуклеотида и пентозой следующего нуклеотида.
Пример соединения аденина и цитозина в одной цепочке молекулы ДНК:
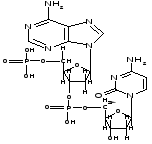
Вторичная структура ДНК имеет форму скрученной лестницы или двойной спирали. Две отдельные цепочки молекулы ДНК соединяются друг с другом посредством относительно слабых водородных связей между азотистыми основаниями нуклеотидов. Такие связи лежат в основе принципа комплементарного спаривания нуклеотидов. При комплементарном спаривании нуклеотидов соблюдаются строгие правила.
Большие (пуриновые) азотистые основания спариваются только с маленькими (пиримидиновыми).
Гуанин в любой молекуле (ДНК и РНК) может спариваться только с цитозином, образуя три водородные связи.
Аденин в молекуле может спариваться с тимином или с урацилом, образуя по две водородные связи. Если аденин спаривается с тимином, то синтезируется или работает молекула ДНК. Если аденин спаривается с урацилом, то синтезируется или работает молекула РНК.
Другие
варианты спаривания нуклеотидов
невозможны. Кратко принцип комплементарного
спаривания азотистых оснований
нуклеотидов записывают так:
Пример соединения двух пар нуклеотидов в молекуле ДНК
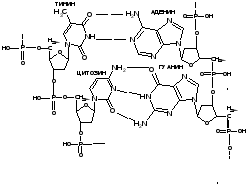
Этот принцип является не только способом связи параллельных цепочек ДНК. Он основа для репликации ДНК и реализации генетической информации (транскрипция при биосинтезе белка).
У некоторых вирусов двухцепочечной может быть РНК.
Связи между параллельными цепочками слабые (водородные), но многочисленные, что делает систему достаточно надежной.
Конкретная последовательность нуклеотидов в цепочках нуклеиновых кислот не случайная. Она передается из поколения в поколение по наследству и определяет генетический код.
Функции ДНК - хранение и реализация генетической информации.
РНК делят на 3 группы.
1. Матричная РНК (м-РНК) и близкая к ней информационная (и-РНК)- это комплементарная копия участка одной из цепочек ДНК. Она получена по принципу комплементарного спаривания азотистых оснований и представляет собой одну полинуклеотидную цепочку из разного количества нуклеотидов (от нескольких десятков до нескольких тысяч).
Функция и-РНК - перенос информации от хранилища (ДНК) к месту реализации, например, на рибосомы для сборки белка. Функционально и-РНК можно разделить на триплетные кодоны (тройки нуклеотидов, кодирующих аминокислоты).
2. Рибосомальные р-РНК, образуют большую и малую субъединицу рибосом. Их функция связана с ролью рибосом, как мест сборки белка.
3. Транспортные т-РНК, имеют сложную пространственную конфигурацию, но на плоскости изображаются в форме клеверного листа. По их главной части - триплетному антикодону - можно различить более 60 разновидностей т-РНК.
Функция т-РНК: перенос аминокислот к рибосомам на встречу с и-РНК для сборки белка.
Липиды - разнородные по строению вещества, как правило, не растворимые в воде и растворимые в органических растворителях.
Наиболее важными для нашего курса биологии являются 3 группы.
1. Нейтральные жиры- это сложные эфиры глицерина и жирных кислот.
Глицерин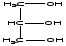
Некоторые жирные кислоты
Насыщенные жирные кислоты
Масляная (С4) СН3-(СН2)2-СООН
Пальмитиновая (С16) СН3-(СН2)14-СООН
Стеариновая (С18) СН3-(СН2)16-СООН
Мононенасыщенные жирные кислоты
Пальмитолеиновая (С16) СН3-(СН2)5-СН=СН-(СН2)7-СООН
Олеиновая (С18) СН3-(СН2)7-СН=СН-(СН2)7-СООН
Полиненасыщенные жирные кислоты
Линолевая (С18) СН3-(СН2)4-СН=СН-СН2-СН=СН-(СН2)7-СООН
(две двойные связи)
Линоленовая (С18) СН3-СН2-СН=СН-СН2-СН=СН-СН2-СН-СН=СН-(СН2)7-СООН
(три двойные связи)
Арахидоновая (С20) СН3-(СН2)4-СН=СН-СН2-СН=СН-СН2-СН-СН=СН-СН2-СН=СН-(СН2)3-СООН
(четыре двойные связи)
Молекула
нейтрального жира может состоять из
различных комбинаций глицерина и жирных
кислот, например, глицерин + линолевая,
пальмитиновая, стеариновая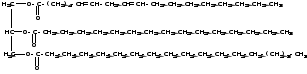
2. Фосфолипиды- триглицериды, в которых одна жирная кислота заменена на фосфорную, что придает глицериновой "головке" молекулы большую гидрофильность, но лишает одного из трех длинных жирно-кислотных гидрофобных "хвостов". Такие молекулы условно изображают в форме двухвостого головастика.
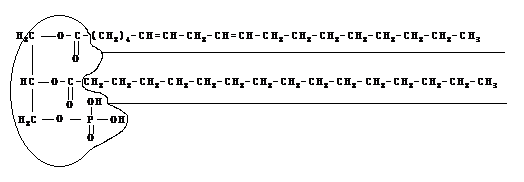
3.
Стероиды - производные
циклопентанпергидрофенантрена.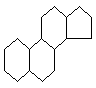
Данная группа соединений дает много полезных соединений (стероидные гормоны, жирные кислоты). В их числе холестерин, который при атеросклерозе портит стенки артерий, а от сердечно- сосудистой патологии умирает более 50 % людей.
Функции липидов.
1. Структурная (строительная). Липиды входят в состав клеточных мембран, например, фосфолипиды - главная (базовая) основа всех клеточных мембран.
2. Защитная. Липиды выполняют функцию механической и термической защиты, например, механическая защита жировой клетчаткой нервных стволов, костей, внутренних органов.
3. Энергетическая. При окислении 1 г жира выделяется энергии (около 40 кДж или 9,3 ккал), почти в 2,5 раза больше, чем при окислении 1 г углевода или белка (17,2 кДж или 4,1 ккал).
4. Регуляторная. Производные циклопентанпергидрофенантрена составляют основу половых гормонов.
Углеводы - соединения содержащие углерод, кислород и водород в пропорции близкой к Cm(H2O)n. В дальнейшем оказалось, что эта пропорция не всегда верна, (например, дезоксирибоза C5H10O4), но название «углеводы» укоренилось.
Три главных класса углеводов: моносахариды, олигосахариды (в т. ч. дисахариды), полисахариды.
1. Моносахариды. Глюкоза, фруктоза, галактоза, сорбит, рибоза, дезоксирибоза
-D-глюкоза -D-фруктоза
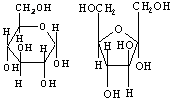
2. Олигосахариды (дисахариды, трисахариды и т. д.): сахароза, лактоза, мальтоза.
дисахарид
сахароза = моносахарид -D-глюкоза
+ моносахарид -D-фруктоза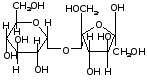
3. Полисахариды - полимеры, мономерами которых являются моносахариды. Например, глюкоза - мономер сразу трех видов гомополисахаридов - крахмала, гликогена, клетчатки.
Функции углеводов.
1. Структурная (строительная). Углеводы входят в состав многих элементов живых организмов, например, клеточная стенка растительной клетки, гликокаликс эпителия кишечника человека.
2. Сигнальная. Углеводно-белковые комплексы (гликопротеиды) образуют рецепторы (см. сигнальная функция белков).
3. Защитная. Гликопротеиды соединительной ткани выполняют функцию химической защиты, противостоят гидролитическим ферментам.
4. Энергетическая. При полном окислении 1 г углеводов выделяется 4,1 ккал или 17,2 кДж энергии.
Эта функция последняя по перечислению, но главная по значению. Углеводы дают человеку более 60% энергии.
Энергетика клетки.
В химических реакциях при образовании связей между простыми молекулами энергия потребляется, а при разрыве выделяется.
В процессе фотосинтеза у зеленых растений энергия солнечного света переходит в энергию химических связей, возникающих между молекулами углекислого газа и воды. Образуется молекула глюкозы: CO2 + H2O + Q (энергия) = C6H12O6.
Глюкоза является главным источником энергии для человека и большинства животных.
Процесс усвоения этой энергии называют " окислительное фосфорилирование". Энергия (Q), выделяющаяся при окислении, сразу используется на фосфорилирование аденозиндифосфорной кислоты (АДФ):
АДФ+Ф+Q (энергия)=АТФ
Получается "универсальная энергетическая валюта" клетки аденозинтрифосфорная кислота (АТФ). Она может в любой момент быть использована на любую полезную организму работу или на согревание.
АТФАДФ+Ф+Q (энергия)
Процесс окисления глюкозы проходит в 2 этапа.
1. Анаэробное (бескислородное) окисление, или гликолиз, происходит на гладкой эндоплазматической сети клетки. В результате этого глюкоза оказывается разорванной на 2 части, а выделившейся энергии достаточно для синтеза двух молекул АТФ.
2. Аэробное (кислородное) окисление. Две части от глюкозы (2 молекулы пировиноградной кислоты) при наличии кислорода продолжают ряд окислительных реакций. Этот этап протекает на митохондриях и приводит к дальнейшему разрыву молекул и выделению энергии.
Результатом второго этапа окисления одной молекулы глюкозы является образование 6 молекул углекислого газа, 6 молекул воды и энергии, которой достаточно для синтеза 36 молекул АТФ.
В качестве субстратов для окисления на втором этапе могут использоваться не только молекулы, полученные из глюкозы, но и молекулы, полученные в результате окисления липидов, белков, спиртов и других энергоемких соединений.
Активная форма уксусной кислоты - А-КоА ( ацетил коэнзим А, или ацетил кофермент А) - это промежуточный продукт окисления всех этих веществ (глюкозы, аминокислот, жирных кислот и других).
А-КоА является точкой пересечения углеводного, белкового и липидного обменов.
При избытке глюкозы и других энергонесущих субстратов организм начинает их депонировать. В этом случае, глюкоза окисляется по обычному пути до молочной и пировиноградной кислоты, затем до А-КоА. Далее, А-КоА становится базой для синтеза молекулы жирных кислот и жиров, которые депонируются в подкожной жировой клетчатке. Наоборот, при недостатке глюкозы, ее синтезируют из белков и жиров через А-КоА (глюконеогенез).
При необходимости могут пополняться и запасы заменимых аминокислот для строительства некоторых белков.
Схема
связи углеводного, липидного, белкового
и энергетического метаболизма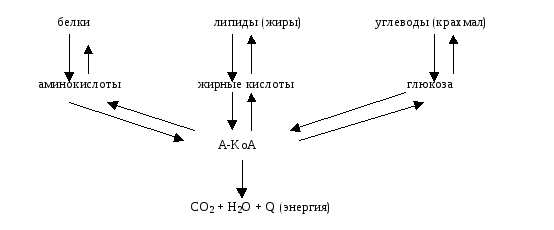
Блок дополнительной информации.
ДНК и Хромосомы.
В ядре диплоидной клетки человека — 46 хромосом.
В пресинтетический (постмитотический) период жизненного цикла делящихся клеток, а также всегда в неделящихся клетках каждая хромосома содержит лишь одну молекулу ДНК. Каждая из этих столь протяженных молекул включает 2 полинуклеотидные цепи, образующие двойную спираль.
Последняя связана с основными (гистоновыми) и кислыми (негистоновыми) белками. Общая длина всех 46 молекул ДНК, находящихся в ядре человеческой клетки, — около 190 см. Эти молекулы существенно различаются по размеру; средняя же длина одной из них, как нетрудно найти, — примерно 4 см.
В свою очередь, каждая цепь — это линейная последовательность нуклеотидов (точнее, дезоксирибонуклеотидов) четырех видов: дАМФ, дГМФ, дЦМФ и дТМФ.
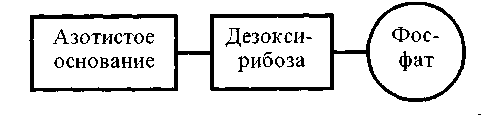
Любой из названных нуклеотидов имеет 3 компонента (рис. 1.).
Исходя из этого, и составляются названия нуклеотидов ДНК: дАМФ —дезоксиаденозинмонофосфат, дГМФ — дезоксигуанозинмонофосфат, дЦМФ — дезоксицитидинмонофосфат, дТМФ — дезокситимидинмонофосфат.
В цепи ДНК нуклеотиды связываются с помощью фосфатных групп (рис. 2).
Рис. 2.Структура ДНК
5' конец З' конец

З' конец 5' конец
Как видно из рис. 2, остов цепей образуется чередующимися остатками пентозы и фосфата. При этом фосфатные остатки расположены между 5'- и 3'-положениями пентоз соседних нуклеотидов. (В нумерации атомов углерода пентозы используются штрихи.)
Азотистые же основания цепей обращены друг к другу, спариваясь по принципу комплементарности: А с Т, Г с Ц.
Принципиальное значение для протекающих на ДНК процессов имеет полярность цепей. На т. н. 5'-конце цепи свободно (5' атом углерода не участвует в образовании межнуклеотидной связи). Соответственно, на З'-конце в образовании межнуклеотидной связи не участвует этот З' атом углерода.
Из представленной схемы также видно, что цепи в молекуле ДНК являются антипараллельными.
При «горизонтальном» изображении ДНК принято располагать ее так, чтобы у верхней цепи слева находился 5'-конец; например:
(5)--АТТГАЦАГГЦ--(3)
(3)--ТААЦТГТЦЦГ--(5)
В двойной спирали на 1 виток ДНК приходится 10 нуклеотидных пар. Длина витка — 3,4 нм.
Примерно 60-80 % хромосомных белков представлены гистонами. Последние обогащены аминокислотами с основными (аргинин, лизин) и гидрофобными (валин и т. п.) радикалами. Благодаря основным радикалам, гистоны взаимодействуют с ДНК, а благодаря гидрофобным радикалам — друг с другом. Эти взаимодействия приводят к образованию нуклеосом. Основа нуклеосомы — глобула из 8 белковых молекул (октамер): она включает по 2 молекулы гистонов четырех видов (Н2А, Н2В, НЗ и Н4). Вокруг одной такой глобулы молекула ДНК делает примерно 2 оборота, что и образует в итоге нуклеосому (рис. 3).
Рис. 3. Три
последовательные нуклеосомы.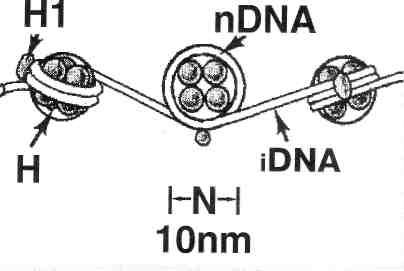
Нуклеосомы расположены на молекуле ДНК не вплотную: между ними имеются линкерные (соединительные) участки (iDNA) длиной 60 н. п. С каждым линкерным участком связана 1 молекула еще одного вида гистонов — HI.
Следовательно, всего в хромосомах встречаются гистоны 5 видов, а период нуклеосомной организации составляет 200 н. п.
Молекула ДНК участвует в образовании очень большого числа нуклеосом (в среднем 600 000). В результате на данном уровне организации каждая хромосома (не считая негистоновых белков) представляет собой длинную нить «бусинок» — нуклеосом диаметром 10 нм. По сравнению с молекулой ДНК, длина нуклеосомной нити примерно в 6,2 раза меньше.
В неделящемся ядре клетки хромосомы не различимы, а воспринимаются все вместе как хроматин.
При этом выделяют гетеро- и эухроматин.
В хроматине к нуклеосомному уровню добавляются последующие уровни укладки хромосомы. Считается, что нуклеосомная нить закручивается в спираль типа соленоида, а та, возможно, образует суперспираль. В этих процессах, видимо, ключевую роль играет гистон HI. В итоге формируется хроматиновая нить диаметром 30 нм.
Хроматиновая нить короче нуклеосомной примерно в 18 раз и короче упакованной в ней молекулы ДНК в 6,2 х 18=100 раз.
В свою очередь, хроматиновые нити образуют петли, которые собираются в розетки, где основания петель крепятся к белкам ядерного матрикса. Наибольшей компактизации хромосомы достигают в процессе деления - митоза (на стадии метафазы). Точная укладка хромосомных нитей при этом неизвестна. По одной из версий, хроматиновая нить многократно складывается по длине хромосомы (рис. 4).
Рис. 4. Укладка нити
ДНК в спирализованной хромосоме.
Дополнительная литература.
1. Грин Н., Стаут У., Тейлор Д. Биология: В 3-х т. Т.1: Пер с англ. / Под ред. Р. Сопера.- М.: Мир, 1993. С.316-319.
2. Мушкамбаров Н.Н., Кузнецов С.Л. Молекулярная биология. М.: 2003 554 с.