

3. Регуляция активности ферментов
Способность к регуляции делает ферменты важными участниками и своеобразными организаторами клеточных процессов в организме человека. Регуляция скорости ферментативных реакций в клетке — основной механизм не только контроля и координации метаболических путей, но и роста и развития клетки, а также ее ответа на изменение окружающей среды.
Существует два основных способа контроля скорости ферментативных реакций:
— Контроль количества фермента.
Количество фермента в клетке определяется соотношением скоростей его синтеза и распада. Этот способ регуляция скорости ферментативной реакции является более медленным процессом (проявляется спустя несколько часов), чем регуляция активности фермента (практически мгновенный ответ).
— Контроль активности фермента.
Активность фермента может регулироваться путем взаимодействия с определенными веществами, изменяющими конформацию активного центра.
Некоторые способы регуляции активности ферментов представлены на рисунке 10.
Регуляция субстратом реакции
Регуляция ферментативной активности, осуществляемая в центре присоединения субстрата, носит название изостерической.
Одним из относительно простых способов регуляции активности ферментов является регуляция с помощью изменения концентрации субстратов реакции. Чем больше в распоряжении фермента имеется молекул веществ, превращения которых он осуществляет, тем выше (до определенных пределов) скорость процесса. При насыщении всех молекул фермента субстратом скорость реакции достигает максимального уровня. В дальнейшем скорость реакции может понизиться по мере исчерпания запасов субстрата и вновь возрасти при их восстановлении.
Слишком большая концентрация субстрата также может понижать скорость ферментативной реакции. Этот феномен носит название субстратного торможения.
В качестве примера субстратного торможения можно привести фермент, расщепляющий биологически активное вещество ацетилхолин - ацетилхолинэстеразу (АХЭ). К активному центру АХЭ субстрат (ацетилхолин) присоединяется двумя концами молекулы одновременно. При увеличении концентрации ацетилхолина с одной молекулой фермента могут одновременно реагировать две молекулы субстрата, но разными концами. В этом случае реакция, суть которой заключается в разрыве сложноэфирной связи в середине молекулы ацетилхолина (с образованием холина и уксусной кислоты), оказывается невозможной, и молекулы ацетилхолинэстеразы, нагруженные субстратом, оказываются тем не менее лишенными активности.
Уменьшение концентрации ацетилхолина в среде приведет к диссоциации неактивного комплекса и снимет торможение. Этот механизм имеет важное физиологическое значение для регуляции концентрации ацетилхолина, который выполняет в нервной системе и мышцах роль медиатора, передающего возбуждение с одной клетки на другую.
Аллостерическая регуляция. Фермент изменяет активность с помощью нековалентно связанного с ним эффектора. Связывание происходит в участке, пространственно удаленном от активного (каталитического) центра (allos - иной). Это связывание вызывает конформационные изменения в молекуле белка, приводящие к изменению определенной геометрии каталитического центра. Активность может увеличиться - это активация фермента, или уменьшиться - это ингибирование.
«Сообщение» о присоединении аллостерического активатора передается посредством конформационных изменений каталитической субъединице, которая становится комплементарной субстрату, и фермент «включается». При удалении активатора фермент вновь переходит в неактивную форму и «выключается». Аллостерическая регуляция является основным способом регуляции метаболических путей.
Метаболические цепи
Обычно ферментативные реакции в клетке организованы в метаболические цепи или циклы, где самая медленная стадия лимитирует скорость всей цепи, то есть последовательности реакций, объединяемых общими субстратами (рис.9).
В таких цепях нередко наблюдается так называемая регуляция по типу обратной связи. Она служит для того, чтобы скорректировать работу цепи с потребностями клетки в конечном продукте. Принцип регуляции заключается в том, что ферменты, стоящие в начале цепи, ингибируются отдаленными метаболитами или конечными продуктами.
Такая регуляция чаще всего происходит по аллостерическому типу, когда молекула регулятора связывается с ферментом в специальном регуляторном центре. Аллостерические ферменты часто выполняют ключевую роль в регуляции обмена веществ, поскольку обладают способностью определять количество важных метаболитов и изменять в соответствии с этим свою активность.
В каждой метаболической цепи есть фермент, который задает скорость всей цепочке реакций. Он называется регуляторным ферментом.
Ферменты, регулирующие скорость метаболических путей:
- обычно действуют на ранних стадиях метаболических путей, в местах ключевых разветвлений метаболических путей;
- катализируют в условиях клетки практически необратимые реакции, протекающие наиболее медленно (ключевые).
|
Метаболическая цепь: А, В, С, D - метаболиты, Е1, Е2, Е3, Е4 - ферменты |
Химическая модификация молекул ферментов (рис.10)
Химическая модификация белков осуществляется за счет присоединения к аминокислотным остаткам в молекуле белка определенных групп: фосфатной группы (при участии протеинкиназ), остатка жирной кислоты (с помощью ацилтрансфераз), углеводных компонентов (гликозил-трансферазы, гликозидазы).
Белки, как правило, имеют лабильную структуру, упаковка которой сильно зависит от свойств химических групп, входящих в состав молекулы. Поэтому присоединение к молекуле белка дополнительных группировок существенно влияет на структуру, а следовательно, и на ферментативную активность молекулы. Такая регуляция носит приспособительный (адаптационный) характер.
Пример. Регуляция активности ферментов путем фосфорилирования-дефосфорилирования. Фермент изменяет активность в результате ковалентной модификации. `
В этом случае фосфатная группа - ОРО32- присоединяется к гидроксильным группам в остатках серина, треонина или тирозина. В зависимости от природы фермента фосфорилирование может его активировать или, наоборот, инактивировать. Реакция присоединения фосфатной группы и ее отщепление катализируют специальные ферменты - протеинкиназы и протеинфосфатазы.
Фосфорилирование - распространенный способ изменить свойства некоторых клеточных белков. Так, при фосфорилировании компонентов цитоскелета (комплекса структурных белков, обеспечивающих поддержание прочности и функционирования клетки) изменяются прочность его взаимодействия с мембраной и форма клеток. Фосфорилирование белков - регуляторов сокращения активирует сократительную реакцию мышцы.
Регуляция с помощью химической модификации белка приводит к долговременным последствиям: модифицированные молекулы сохраняют свои функции измененными до тех пор, пока специальные ферменты не отщепят модифицирующую белок химическую группу и не вернут его в исходное состояние.
Регуляция путем белок-белковых взаимодействий (ассоциации-диссоциации субъединиц в олигомерном ферменте). (рис.10) Например, фермент протеинкиназа в неактивной форме построена как тетрамер R2C2 (R и С - разные субъединицы). Активная протеинкиназа представляет собой субъединицу С, для освобождения которой необходима диссоциация комплекса. Активация фермента происходит при участии цAMP, который способен присоединиться к субъединице R, после чего изменяется конформация, комплементарность субъединиц R и С и происходит диссоциация комплекса: R2C2 + 2cАМР 2С + 2(R -цАМР)
Протеинкиназа фосфорилирует соответствующие ферменты, изменяет их активность и, следовательно, скорость метаболизма в клетке.
Активация ферментов путем частичного протеолиза
Чачтичный протеолиз-разрушение белковой структкры до аминокислотных остатков,для того.чтобы слизистая пожелудочной не разрушилась от трипсина
. Некоторые ферменты синтезируются первоначально неактивными и лишь после секреции из клетки переходят в активную форму. Неактивный предшественник называется проферментом. Активация профермента включает модификацию первичной структуры с одновременным изменением конформации. Например, трипсиноген, синтезированный в поджелудочной железе, затем в кишечнике превращается в трипсин путем удаления фрагмента с N-конца гексапептида. Расщепление определенных пептидных связей «запускает» новые взаимодействия R-групп по всей молекуле, приводя к новой конформации, в которой R-группы активного центра занимают оптимальное положение для катализа (рис.10).
Роль липидного окружения.
Изменение вязкости микроокружения белковых молекул управляет взаимодействием между белками в олигомерных комплексах и регулирует активность мембраносвязанных ферментов.. Этот тип регуляции, который обнаружен в случае многих мембранных белков, обеспечивает тонкую настройку их работы на сиюминутные потребности клетки.
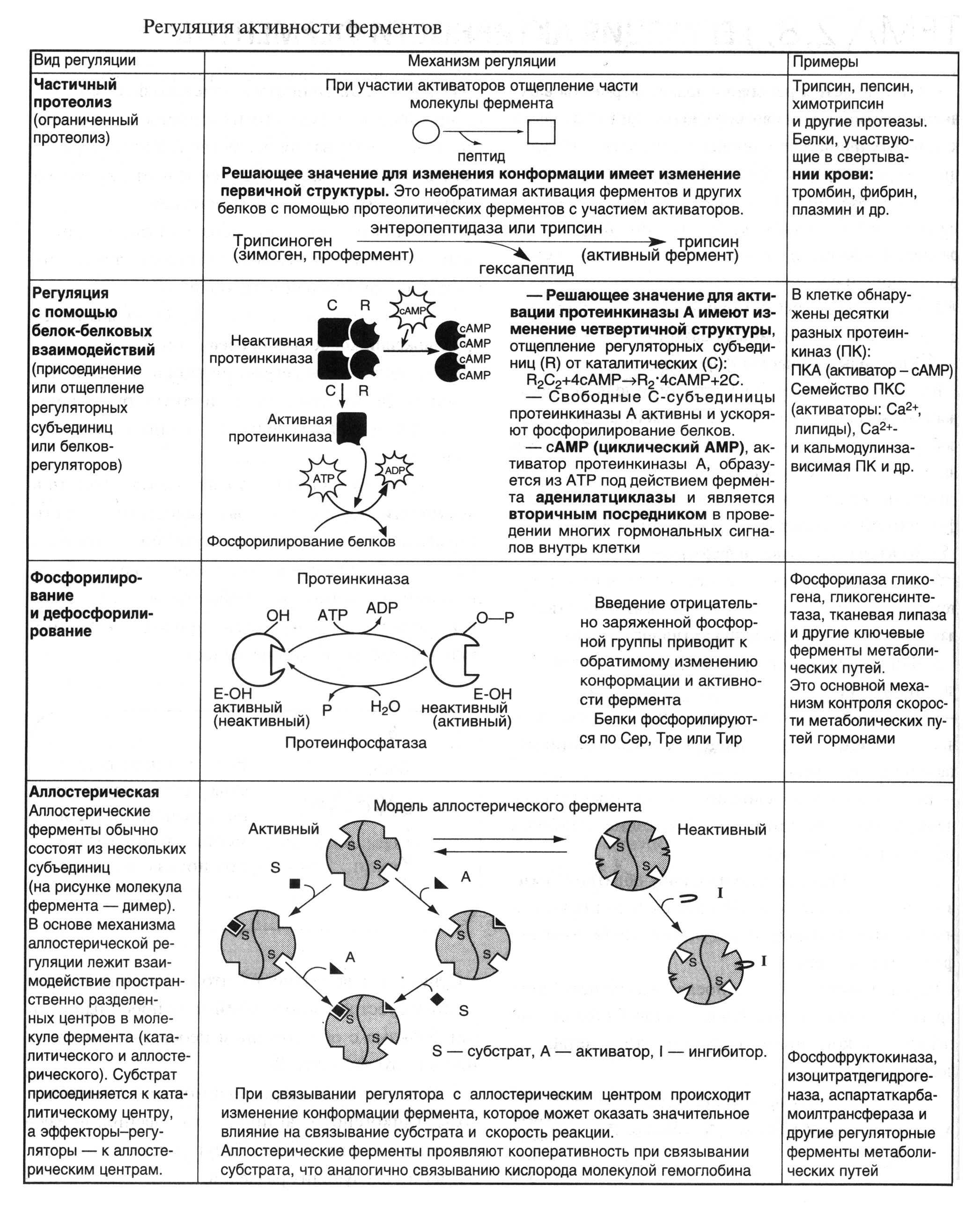
Рис.10