

Типы рнк:
Матричная (информационная) РНК — РНК, которая служит посредником при передаче информации, закодированной в ДНК к рибосомам, молекулярным машинам, синтезирующим белки живого организма. Кодирующая последовательность мРНК определяет последовательность аминокислот полипептидной цепи белка. Транспортные (тРНК).Они переносят специфические аминокислоты в место синтеза пептидной связи в рибосоме. Каждая тРНК содержит участок для присоединения аминокислоты и антикодон для узнавания и присоединения к кодонам мРНК. Рибосомальные РНК (рРНК) — каталитическая составляющая рибосом. тмРНК - обнаружен во многих бактериях и пластидах. При остановке рибосомы на дефектных мРНК без стоп-кодонов тмРНК присоединяет небольшой пептид, направляющий белок на деградацию. Малые интерферирующие РНК (миРНК) часто образуются в результате расщепления вирусных РНК, но существуют и эндогенные клеточные миРНК. Малые ядерные РНК.
Концепция «мир РНК». В живых организмах практически все процессы происходят в основном благодаря ферментам белковой природы. Белки, однако, не могут самореплицироваться и синтезируются в клетке de novo на основании информации, заложенной в ДНК. Но и удвоение ДНК происходит только благодаря участию белков и РНК. Образуется замкнутый круг, из-за которого, в рамках теории самозарождения жизни приходилось признать необходимость не только абиогенного синтеза обоих классов молекул, но и спонтанного возникновения сложной системы их взаимосвязи. В начале 1980-х годов в лаборатории Т. Чека и С. Олтмана в США была открыта каталитическая способность РНК. По аналогии с ферментами (англ. enzyme) РНК-катализаторы были названы рибозимами, за их открытие Томасу Чеку в 1989 году была присуждена Нобелевская премия по химии. Более того, оказалось, что активный центр рибосом содержит большое количество рРНК. Также РНК способны создавать двойную цепочку и самореплицироваться. Таким образом, РНК могли существовать полностью автономно, катализируя «метаболические» реакции, например, синтеза новых рибонуклеотидов и самовоспроизводясь, сохраняя из «поколения» в «поколение» каталитические свойства. Накопление случайных мутаций привело к появлению РНК, катализирующих синтез определённых белков, являющихся более эффективным катализатором, в связи с чем эти мутации закреплялись в ходе естественного отбора. С другой стороны возникли специализированные хранилища генетической информации — ДНК. РНК сохранилась между ними как посредник.
-
Макромолекулярная структура рнк. Формирование пространственной структуры у тРнк и рРнк.
Первичная структура РНК. 1965 году Р.Холли установили нуклеотидную последовательность первичной молекулы РНК.
Вторичная
структура - совокупность
всех непрерывных «стеблей» (участков
спаривания) в пределах одной молекулы
первичной конформации РНК. Основной
компонент вторичных структур -
относительно короткие двойные спирали,
образованные комплементарными участками
одной и той же цепи и перемежающиеся ее
однотяжевыми сегментами. Полирибонулкеотидные
цепи антипараллельны, а сами двойные
спирали находятся в А-форме: в них имеются
дефекты в виде неспаренных нуклеотидных
остатков или петель. Стабильность
двутяжевых районов поддерживается
комплементарными и межплоскостными
взаимодействиями оснований. В однотяжевых
участках – стэкинг-взаимодействия,
вследствии принимают форму однотяжевой
спирали. Одноцепочечные (неспаренные)
участки молекулы называют «петлями».
Петли бывают трех типов: шпилечные,
внутренние и мультипетли. 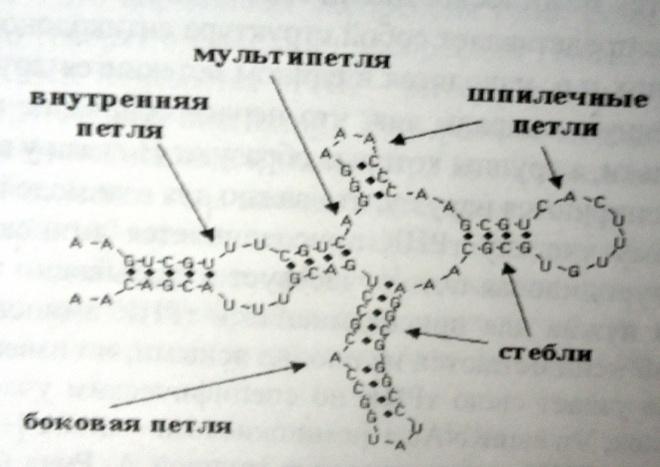
Третичная структура. Возникает за счет взаимодействия элементов вторичной структуры. 1.Элементы вторичной структуры РНК располагаются друг относительно друга так, чтобы обеспечить максимальный стэкинг оснований в макромолекуле в целом. 2. Контакты между отдельными элементами вторичной структуры осущ за счет несколькх типов внутримолекулярных взаимодействий: а)за счет образования дополнительных не уотсон-криковских пар оснований между нуклеотидными остатками удаленных друг от друга; б)за счет дополнительных стэкинг-взаимодействий после внедрения оснований одного участка между двумя соседними основаниями другого однотяжевого участка; в) за счет образования дополнительных водородных связей между 2`ОН-группами остатков рибозы и оснований. 3.Третичная структура РНК стабилизирована ионами двухвалентных металлов, которые связываются не только с фосфатными группами, но и с основаниями.
рРНК
прокариот
и эукариот
отличаются друг от друга, хотя и
обнаруживают значительное сходство
участков последовательностей.
Высокомолекулярные рРНК являются
структурной основой для формирования
рибонуклепротеинового тяжа, который
складывается в пространстве и дает
начало 30-40s
и 50-60-субъединицам рибосомы. Низкомолекулярная
5SpРНК
в комплексе с рибосомными белками
формирует третью субъединицу, выступает
в роли посредника между пептидилтрансферазным
центром и EF-G-связывающими
доменами. 
