

Синтез и механизм действия внутриклеточных гормонов
1.Циклические нуклеотиды
а) ц-3′,5′ АМФ
Мембрану клетки пронизывает сложная белковая система – аденилатциклаза, включающая три субъединицы: рецепторную (R), G-белок (G), каталитическую (С). (Рис. 1).
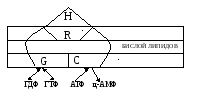
Рис.1. Схема работы аденилатциклазы в клеточной мембране.
Е


 сли
гормон (Н) комплементарен рецептору
(R), то они комплексируются вследствие
образования между ними слабых связей;
при этом несколько меняется структура
R-субъединицы, что передаётся G-белку,
который активируется и вызывает изменения
в балансе системы ГДФ ГТФ.
Последнее стимулирует работу С-субъединицы.
Этот катализатор обусловливает циклизацию
АТФ: АТФц-3′,5′
АМФ.
сли
гормон (Н) комплементарен рецептору
(R), то они комплексируются вследствие
образования между ними слабых связей;
при этом несколько меняется структура
R-субъединицы, что передаётся G-белку,
который активируется и вызывает изменения
в балансе системы ГДФ ГТФ.
Последнее стимулирует работу С-субъединицы.
Этот катализатор обусловливает циклизацию
АТФ: АТФц-3′,5′
АМФ.
ФФ
Получившееся соединение служит активатором протеинкиназы А:
цАМФ
Б
 елок
+ АТФ Белок ~ Ф + АДФ
елок
+ АТФ Белок ~ Ф + АДФ
Протеинкиназа А
Фосфорилирование протеина меняет его способность выполнять функции. Если фосфат присоединяется к ферменту, то варьирует его активность (например, фосфорилирование фосфорилазы гликогена стимулирует гликогенолиз, а подобная модификация гликогенсинтазы угнетает образование данного полисахарида). Когда фосфат включается в белки мембран, то меняется их заряд и упаковка, что сказывается на проницаемости, текучести и других свойствах данных органоидов. Фосфорилирование ядерных протеинов изменяет скорость экспрессии генов, различные геномные процессы.
цАМФ – служит вторичным посредником для адреналина, глюкагона, кальцитонина, АКТГ, ТТГ и других гормонов.
Гашение данного сигнала обеспечивается фосфодиэстеразой (ФДЭ):
 цАМФ + Н2О
5′-АМФ
цАМФ + Н2О
5′-АМФ
ФДЭ
б) ц-3′,5′ ГМФ
Механизм работы данного циклического нуклеотида аналогичен. Отличается тем, что в гуанилатциклазе отсутствует G-белок и образующийся цГМФ активирует иной вид ферментов (протеинкиназы G), изменяя функции других протеинов, в первую очередь, отвечающих за работу ионных каналов. Через гуанилатциклазную систему действуют натрийуретический предсердный пептид, монооксид азота, монооксид углерода.
2. Активация протеинкиназ без предварительного образования вторичного посредника.
Локализующаяся в мембране клетки тирозинкиназа после комплексирования своей рецепторной субъединицы с гормоном активируется; при этом преобразуется её каталитическое звено, начинается фосфорилирование тирозина, содержащегося в близлежаших протеинах, что сказывается на выполнении ими своих функций. Примером гормонов, действующих на тирозинкиназу, могут служить инсулин, цитокины.
3,4. Последствия активации фосфолипазы С.
Этот фермент, расположенный в мембранах, после своей активации гормонами (АДГ, окситоцином) катализирует частичный гидролиз инозитолфосфатидов с последующим фосфорилированием продуктов:
Фосфолипаза С
И
 нозитолфосфатидыДАГ
+ И3Ф
нозитолфосфатидыДАГ
+ И3Ф
АТФ Фосфокиназы
Диацилглицеролы (ДАГ), оставаясь в плазмолемме, служат активаторами протеинкиназ С, фосфорилирующих специфические белки с последующими изменениями их функций. Инозитолтрифосфаты (И3Ф) обладают сильным отрицательным зарядом и притягивают в клетку ионы Са++, которые, связываясь с различными полипептидами, активно влияют на их деятельность (изменяя активность ферментов, свойства протеинов мембран, поведенческую память, обучение и др.).