

Последние и будут (возможно, при вскармливании или иным путем)
прививаться детям. Не исключено, что эти поликультуры будут собираться у наиболее здоровых матерей. Следует также выявить, будут ли оптимальные поликультуры в различных странах идентичны или должны различаться в связи с климатическими и другими особенностями жизни различных групп людей.
3.6. Элементные диеты и две теории питания
Напомним, что идея элементного (мономерного) питания, возникшая в конце XIX-начале XX в. и вытекающая из теории сбалансированного
питания, заключалась в том, что нашу традиционную пищу можно и целесообразно заменить легкоусвояемыми мономерами, имитирующими смесь
продуктов |
питания - |
белков, углеводов, жиров и др. Считалось, что |
такая пища вполне удовлетворит потребности организма. |
||
Однако |
с позиций |
теории адекватного питания элементные диеты |
дефектны по многим причинам и, в частности, потому, что нарушают свойства и соотношение нутритивного (трофического) и токсического
потоков вследствие выпадения защитных функций мембранного |
пищеварения |
и изменения эндоэкологии. Действительно, у моногастричных |
организмов |
(в том числе у человека) питание бактерий построено на использовании
преимущественно неутилизируемых или медленно утилизируемых
макроорганизмами компонентов пищи. Мембранное пищеварение, реализуемое ферментами, локализованными в недоступной бактериям щеточной кайме,
препятствует поглощению ими нутриентов и обеспечивает стерильность процесса (Уголев, 1967, 1985). Такую стерильность можно рассматривать как приспособление макроорганизма к сосуществованию с кишечной бактериальной флорой и как фактор, обеспечивающий преимущественное поглощение нутриентов макроорганизмом. Если же пища вводится в
организм в виде мономеров, то мембранное пищеварение как защитный механизм не функционирует. В этом случае бактерии оказываются в чрезвычайно благоприятных условиях для их размножения в результате
избытка легкоусвояемых элементов в полости тонкой кишки. Это приводит
к нарушению эндоэкологии, к увеличению потока токсических веществ и к потере макроорганизмом ряда веществ, в том числе необходимых. При изучении эффектов мономерного питания нами, а затем многими другими исследователями зарегистрированы дисбактериозы и дополнительное
дезаминирование аминокислот (обзор: Уголев, 1985).
Далее, имеются многочисленные сведения, касающиеся более быстрого
(или идентичного) всасывания в тонкой кишке человека и животных олиго-
и полисахаридов по сравнению с составляющими их свободными
86
моносахаридами. Существует также множество данных о больших скоростях всасывания пептидов по сравнению с составляющими их свободными аминокислотами. Особенно интересно, что всасывание ферментных гидролизатов белков, состоящих преимущественно из олигопептидов,
происходит быстрее, чем эквивалентной смеси аминокислот, симулирующих эти белки (обзоры: Химические и физиологические проблемы..., 1975, 1976; Silk, 1977; Gardner, 1984, 1988, и др.).
При специальном исследовании аккумуляции углеводов различной степени полимеризации в слизистой оболочке тонкой кишки крыс нами было
обнаружено, что транспорт свободной глюкозы не зависит от состава
диет, т.е. является стабильным процессом, в отличие от этого транспорт
глюкозы, освобождающейся при гидролизе мальтозы и |
крахмала, |
|||
значительно меняется и зависит от |
состава рациона (обзор: |
Уголев, |
||
1972). |
Сходные результаты получены |
в Институте биологии |
АН |
Латвии |
(Рига) |
на цыплятах (обзор: Кушак, 1983). |
|
|
|
Феномен более быстрого транспорта мономеров, образующихся при гидролизе олиго- и полимеров, чем свободных мономеров, может найти объяснение в свете концепции мембранного пищеварения. Эта концепция предполагает пространственную и структурнофункциональную интеграцию заключительных этапов гидролиза и начальных этапов всасывания. Многими принимается, что интеграция реализуется за счет ферментно-транспортных комплексов мембраны, в которых интегрированы один или несколько ферментов и одна или несколько транспортных систем, реализующих трансмембранный перенос продуктов гидролиза. Идея такого комплекса
сохраняет |
свое значение как |
для концепции мобильного переносчика, так |
и для |
гипотезы канала |
(Уголев, 1972, 1985, 1987а; Membrane |
digestion..., 1989). В ферментно-транспортном комплексе продукты гидролиза должны непосредственно передаваться с активного центра фермента на вход в транспортную систему мембраны с минимальным рассеиванием в водной фазе, а транспортная система не должна взаимодействовать с мономерами, присутствующими в водной среде
(рис.3.3).
87
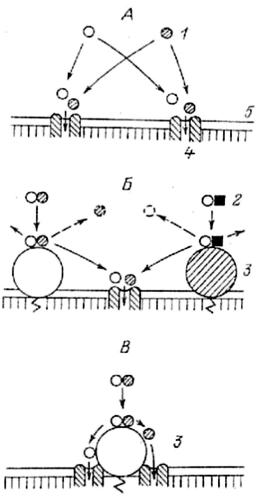
Рис.3.3. Роль ферментно-транспортных комплексов в предотвращении конкуренции между мономерами на стадии всасывания
(по: Уголев, 1985).
А - конкуренция между мономерами за общий вход в транспортную систему; Б - конкуренция между конечными продуктами гидролиза за общий вход в транспортную систему; В - ферментно-транспортный комплекс: передача конечных продуктов гидролиза с фермента на вход в транспортную систему (без конкуренции). 1 - мономер; 2 - димер; 3 - фермент; 4 - транспортная система; 5 - мембрана.
Существуют отрицательные последствия длительного применения
мономерных диет, охарактеризованные в ряде специальных сводок, в том числе в сводках «Химические и физиологические проблемы... »,
опубликованных в Риге в 1975 и 1976 гг. К таким последствиям
относятся, в частности, замедление роста и уменьшение массы тела
животных, увеличение экскреции аммиака, уменьшение экскреции
электролитов, развитие гемолитической анемии и т.д. Недавно показано
также, что при длительном использовании элементных диет происходит
трансформация некоторых вводимых в организм лекарственных препаратов в
токсические формы (Interaction of elemental diets..., 1988). Кроме
того, мономерные диеты приводят к снижению функциональной нагрузки на
ферментные системы желудочно-кишечного тракта, что сопровождается
нарушением синтеза ряда ферментов, которые необходимы для нормальной
деятельности организма. Далее, из-за высокой осмотической активности
элементных диет нарушается распределение жидкости между кровью и
энтеральной средой в результате перехода жидкости из крови в кишечник.
Однако при некоторых формах заболеваний и в определенных условиях
элементные и без балластные диеты могут быть весьма полезны. В
88
частности, при врожденных и приобретенных дефектах ферментных систем
тонкой кишки наиболее целесообразно исключать из пищи те вещества
(например, лактозу, сахарозу и др.), гидролиз которых нарушен.
Элементные диеты могут быть использованы при различных экстремальных воздействиях, вызывающих нарушения деятельности желудочно-кишечного тракта. При этом неадекватность, например имитаций белка набором определенных аминокислот, проявляется не сразу, а через определенный временной интервал, в течение которого эти аминокислоты могут служить полноценной заменой белка. Вероятно, отрицательные последствия элементных диет связаны с изменением бактериального состава или по крайней мере с изменением свойств бактериальной популяции кишечника.
Элементные, или мономерные, диеты важны при патологических состояниях, при которых наблюдается репрессия синтеза и включения в
состав мембраны кишечных клеток ферментов, осуществляющих
заключительные этапы пищеварения. В этом случае всасывания аминокислот и гексоз, входящих в состав олигомеров, не происходит. Такие феномены могут наблюдаться, в частности, под влиянием стрессорных факторов.
Тогда аминокислоты могут быть использованы для поддержания удовлетворительного азотистого баланса, отрицательного при стрессе,
для которого характерна потеря белков. Такой отрицательный азотистый баланс возникает за счет глюконеогенеза. Мы получили результаты,
расширяющие классические представления о происхождении отрицательного
азотистого баланса, которые опубликованы в 1972 г. (Уголев, 1972).
Нами обнаружено, что при стрессе происходит снижение уровня дисахаридазных и особенно пептидазных активностей тонкой кишки за счет торможения включения ферментов в состав апикальной мембраны кишечных клеток, что приводит к ослаблению ассимиляции углеводов и главным образом белков. Таким образом, при стрессе отрицательный азотистый баланс обусловлен не только разрушением, но и недостаточным поступлением аминокислот во внутреннюю среду организма. Следовательно,
при различных видах стресса существует эффективный путь коррекции белкового обмена за счет введения в рацион вместо белков, которые не
усваиваются, |
имитирующих |
эти |
белки |
аминокислотных |
смесей. |
Использование |
подобных смесей |
целесообразно также в |
аварийных |
||
ситуациях, при чрезмерных физических нагрузках (например, в условиях
экспедиций), при спортивных соревнованиях, при хирургических вмешательствах, а также в пред- и послеоперационные периоды, в
условиях недостатка белка, при голоде, при высокой или низкой
температуре окружающей среды, при ряде заболеваний и т.д., т.е. в
случаях, которые сопровождаются возникновением стресса - общей для
89