

2. Світловий мікроскоп і його будова. Правила роботи з
імерсійною системою. Фазово-контрастна, люмінесцентна,
електронна мікроскопія

Мікроорганізми дуже малі за своїми розмірами, так що побачити їх неозброєним оком неможливо. Для їх виявлення й вивчення використовують складні прилади - мікроскопи. Сучасна промисловість випускає багато видів мікроскопів залежно від їх призначення
Мікроскоп складається з механічної, оптичної й освітлюваної частин. До механічної частини входять: штатив, тубус, "револьвер", предметний столик, макро- й мікрогвинт. До оптичної частини відносять Рис. і. Мікроскоп об'єктиви й окуляри. До освітлювальної частини - МБР-1 дзеркало й конденсор (рис. 1).
У верхній частині штатива є тубус у який вставляється окуляр, а знизу він має "револьвер"- пластину, що обертається по колу, в отвори якої вгвинчені 3-4 об'єктиви. Обертаючи "револьвер", можна встановити будь-який об'єктив під отвір тубуса. Останній пересувається за допомогою макро- й мікрометричного гвинтів. Для грубого наведення тубуса користуються макрогвинтом. Більш точно це роблять за допомогою мікрогвинта, що дає чітке зображення досліджуваних об'єктів. Мікрометричний гвинт є однією з найбільш тендітних частин мікроскопа й вимагає обережного поводження з ним.
Предметний столик має круглу або прямокутну форму, в його центрі знаходиться отвір, над яким вміщують предметне скло з препаратом. У більш досконалих мікроскопах є дуже зручні предметні столики, які за допомогою спеціальних пристроїв переміщують предметне скло у двох взаємно перпендикулярних напрямах.
Найціннішою частиною мікроскопа є об'єктиви, які складаються з кількох лінз у загальній металевій оправі. Об'єктиви поділяються на сухі (х8, х40) та імерсійні (х90, х120). Сухими називають такі об'єктиви, між фронтальною лінзою і предметним склом яких
знаходиться повітря. При цьому, в зв'язку з різницею показників заломлення "скла й повітря (відповідно 1,52 і 1,0), частина світлових променів не потрапляє в око мікроскопіста. Імерсійними називають такі об'єктиви, між фронтальною лінзою яких і досліджуваним об'єктом знаходиться кедрова, персикова олія, "імерсіол", кофіцієнт заломлення яких такий самий, як і в скла. При дослідженні мікроорганізмів користуються переважно імерсійними об'єктивами, які часто називають імерсійною системою.
Окуляр складається з двох лінз і тільки збільшує зображення, яке виходить від об'єктива. Існують окуляри з такими збільшеннями: х7, хІО, х15. Ці цифри позначені на них.
Загальне збільшення мікроскопа дорівнює добуткові збільшення об'єктива на збільшення окуляра. Наприклад, комбінація імерсійного об'єктива з показником збільшення х90 й окуляра хІО дає збільшення у 900 разів. Максимальне збільшення сучасних мікроскопів досягає 2000-3000. Однак, якість мікроскопа визначається не тільки ступенем збільшення об'єкта, а й властивою йому, так званою роздільною здатністю. Це та найменша відстань між двома точками, зображення яких виразне під мікроскопом. Роздільна здатність сучасних імерсійних об'єктивів знаходиться в межах 0,2 мкм.
Освітлювальний апарат знаходиться під предметним столиком і складається із дзеркала та конденсора з діафрагмою. Дзеркало спрямовує пучок світла в конденсор,'а через нього - в об'єктив мікроскопа. Один бік дзеркала плоский, другий - вігнутий. При мікроскопуванні з конденсором необхідно користуватись лише плоским дзеркалом. Конденсор Аббе складається з системи лінз для збирання пучка променів в одній точці (фокус) яка знаходиться в площині досліджуваного препарату. При роботі з денним освітленням конденсор потрібно підіймати до рівня предметного столика, з штучним - опускати доти, поки зображення джерела світла не з'явиться в площині препарату. При дослідженні незабарвлених препаратів конденсор також опускають.
Об'єм світла відповідно до потреб дослідження регулюється діафрагмою, яка знаходиться під конденсором. Вона може звужуватись і розширюватись подібно до зіниці ока (звідси назва іріс-діафрагма). Забарвлені препарати розглядають при відкритій, а незабарвлені -при звуженій діафрагмі.
Сучасні мікроскопи мають ряд удосконалень, завдяки яким покращується зображення та розширюються межі видимості. Перше
26
27
на використанні явища флуоресценції. Так як більшість хвороботвор них мікроорганізмів не мають первинної (власної) люмінесценції, їх спочатку фарбують слабкими розчинами спеціальних барвників (флу- орохромів), які зв'язуються певними структурами клітин. Із них частіше використовують акридин оранжевий, аурамін, корифосфін, флуоресцеїн. Наприк лад, при забарвлюванні дифтерійних паличок корифосфіном, вони набувають жовто-зеле ного світіння в ультрафіолетовому світлі, збудник туберкульозу при забарвлюванні аурамін-родаміном - золотисно-оранжевого. Метод люмінесцентної мікроскопії набагато Рис. 3. Мікроскоп чутливіший порівняно з іншими мікроскопіч-
люмшесцснтний ними дослідженнями. Він дозволяє виявити в
матеріалі таку малу кількість збудника, яку іншими методами не знаходять (рис. 3).
Люмінесцентну мікроскопію використовують для виявлення антигенів і антитіл (метод імунофлуоресценції). З її допомогою можна побачити мікроби, які містять певні антигени. Для їх виявлення необхідно мати специфічні люмінесцентні сироватки, які викликають флуоресценцію саме даного антигена. Порівняно із звичайними методами світлової мікроскопії, використання люмінесцентних мікроскопів має ряд переваг: кольорове зображення, більша контрастність, можливість досліджувати як живі, так і вбиті мікроорганізми. Цей метод можна використати для експрес-діагностики багатьох бактерійних і вірусних захворювань.
Електронна мікроскопія. Можливості світлового мікроскопа обмежені не якістю лінз, а великою довжиною світлових хвиль (2,8-0,29 мкм). Частинки менших розмірів знаходяться за межами роздільної здатності світлових мікроскопів. Для дослідження таких дрібних об'єктів використовують електронний мікроскоп - високовольтний вакуумний прилад, у якому збільшене зображення отримують за допомогою потоку електронів. Він має високу роздільну здатність і може давати збільшення від 20 тис до 5 млн разів. За принципом дії розрізняють просвічуючі (трансмісивні), скануючі (растрові) й комбіновані електронні мікроскопи.
Принципова схема просвічуючого електронного мікроскопа мало чим відрізняється від схеми світлового. В електронному мікроскопі замість світла використовують потік електронів, джерелом яких є
вольфрамова нитка, що нагрівається електричним струмом (електронна пушка). Роль лінз виконує кругове електромагнітне поле. Пучки електронів, проходячи через досліджуваний об'єкт, відхиляються під різними кутами залежно від неоднакової товщини й щільності препарату. Це й забезпечує контрастність зображення.
Досліджуваний об'єкт спочатку зафіксовують спеціальними фіксаторами, потім наносять на надзвичайно тонку колодієву або целюлозну плівку, вміщену на спеціальну сіточку-підкладку. Для визначення деталей структури бактерій чи вірусів застосовують метод напилення або негативного контрастування. При напиленні на поверхню препарату під певним кутом наносять тонким шаром у вакуумі різні важкі метали, хром, золото, паладій. Розпорошені частинки металу осідають на піднесених чи заглиблених ділянках бактерій або вірусів. При дослідженні таких препаратів деталі їх структури проявляються рельєфно і контрастно. Негативне контрастування зводиться до нанесення на препарат розчинів з атомами важких металів, наприклад, фосфорно-вольфрамової кислоти. Осідаючи навколо білкових частинок досліджуваного об'єкту й заповнюючи всі проміжки між ними, атоми важких металів "забарвлю-ють"фон, на якому виступають найменші деталі будови мікроорганізмів.

Широко також використовують ультратонкі зрізи клітин, бактерій і вірусів, що дає змогу вивчити їх структуру на субклітинному й молекулярному рівнях.
Сучасна українська й зарубіжна промисловість випускає багато моделей електронних мікроскопів (рис. 4), які мають величезні можливості для вивчення мікроскопічного світу.
Методи електронної мікроскопії привели до великих успіхів у цитології, генетиці, бактеріології І, особливо, ВІруСОЛОГІЇ. УСПІШНО роЗВИВаЄТЬСЯ Рис. 4. Мікроскоп
імунна електронна мікроскопія, яка дає змогу електронний визначити родову належність вірусів, що використовується для експрес-діагностики багатьох вірусних інфекцій.
Практична робота
Оволодіти правилами роботи з імерсійною системою мікроскопа.
Вивчити під мікроскопом морфологію бактерій в готових препа-ратах-мазках і замалювати їх .
зо
■
3. Ознайомитись з правилами роботи в мікробіологічній лабораторії. Питання. для самоконтролю
Що вивчає наука мікробіологія?
Які основні досягнення і завдання мікробіологічної науки?
Основні періоди розвитку мікробіологи?
Значення робіт А. Левенгука і Е. Дженнера?
Вклад Л. Пастера в розвиток і становлення медичної мікробіологи.
Діяльність Р. Коха - творця бактеріологічної техніки.
Значення робіт І.І. Мечнікова для розвитку вітчизняної і світової мікробіологічної науки.
Д.И. Івановський - основоположник науки про віруси.
Вклад українських учених у розвиток мікробіологи.
Які правила мікроскопування з імерсійною системою?
32
Розділ 2. МОРФОЛОГІЯ МІКРООРГАНІЗМІВ
Мікроорганізми - загальна збірна назва для найдрібніших організмів Це наистародавніші живі істоти на Землі. До них належать бактерії віруси, гриби, найпростіші. Бактерії разом із водоростями складають царство прокаріотів; віруси - неклітинні форми життя, які мають власний геном і відтворюються тільки в клітинах; найпростіші та гриби - одно- або багатоклітинні еукаріотичні організми.
Класифікація й номенклатура бактерій
Одна з перших спроб наукової класифікації бактерій належить датському зоологу О. Мюллеру, який ще у,XVIII ст. виділив два роди - Мопаз і УіЬгіо. Пізніше німецький біолог Е. Геккель запропонував виділити мікроби в окреме царство Ргоїізіа (ргоїоз - найпростіший). Воно охоплювало переважно одноклітинні мікроорганізми Подальші дослідження привели до поділу цього царства на вищі (водорості, гриби, найпростіші) і нижчі (бактерії, ціанобактерії) протисти іакии підхід сприяв чіткому поділу живого світу на прокаріотичні та еукаріотичні живі системи.
Сучасна систематика (таксономія) бактерій - наука про їх розподіл за певними групами (таксонами). Для їх характеристики враховують різноманітні властивості: морфологічні ознаки, здатність спожива™ атмосферний кисень, шляхи одержання енергії, оптимальна температура росту, рН середовища, здатність засвоювати певні речовини, наявність включень, склад клітинної стінки, вміст основ ДШ\., екологічна ніша тощо.
Для назви мікроорганізмів використовують подвійну номенклатуру К. Лшнея. Перше слово вказує на рід і пишеться з великої літери Воно походить від прізвища вченого, який відкрив і вивчав даний мікроорганізм, або характеризує якусь морфологічну ознаку Друге слово означає вид, пишеться з малої літери і пов'язане з назвою хвороби прізвищем автора та ін. Так, стафілокок золотистий має
гі?,3 к Р Т ССи3 аИГЄи5' 3буДНИК тУберкульозу - МусоЬасіе-гшт ШЬегсиІозіз, кишкова паличка - Езспегіспіа соїі
33
Найбільше визнання серед мікробіологів отримала класифікація мікроорганізмів, яка подана у Визначнику бактерій Д. Бергі (Вещеу'з Мапиаі Зузїетаііс Васїегіо1о§у). КУ
Морфологія і
ультраструктура бактерій
Відповідно до Міжнародного кодексу номенклатури бактерій мікробіологи використовують такі таксономічні критерії систематики: вид - рід - родина - порядок - клас - відділ - царство.
Відповідно до неї царство Ргосагуоіае поділено на 4 відділи за особливостями будови клітинної стінки, відношенням до фарбування за методом Грама та ін.: Сгасіїісиіез (дгасіїіз - тонкий, сшіз - шкіра) -грамнегативні бактерії, Рігтісиіез (іігтиз - міцний) - грампозитивні бактерії, Тепегісиіез (іепег - м'який, ніжний) - мікроби, які не мають клітинної стінки, Мепсіозісиіез (тепйозиз - помилковий) - представники мікробного світу, що мають нетиповий пептидоглікан.
Згідно з цим визначником усі мікроорганізми поділені на 33 групи за ознаками, які винесено у назву групи: 1 - спірохети, 4 - грамнегативні аеробні палички й коки, 12 - грампозитивні коки, 13 - грампозитивні палички й бактерії, що утворюють спори, тощо. Всередині груп поділ мікробів відбувається на порядки, родини, роди, види.
Однак генетичні механізми, що лежать в основі мінливості, здатні забезпечувати тільки відносну стабільність ознак в межах одного ви ду, тому введено поняття про варіанти (типи) бактерій, які за деякими особливостями відрізняються від стандартних видів: морфовари (за морфологічними ознаками), біовари (за біологічними), ферменто-вари (за ферментативними), фаговари (за чутливістю до бактеріофагів), серовари (за антигенними властивостями), ековари (за екологіч ними нішами помешкання), патовари (за патогенністю для лаборатор ■ них тварин).
Існує таке важливе поняття як "штам" - сукупність мікробних особин одного виду, які виділено з різних джерел (організму людини, тварини, зовнішнього середовища), або з одного джерела, однак у різний час.
У популяційну мікробіологію введено поняття "клон" - нащадки однієї мікробної клітини.
34
Зовнішній вид мікроорганізмів напрочуд різноманітний. За формою бактерії поділяють на кокоподібні, паличкоподібні, звивисті та ниткоподібні.
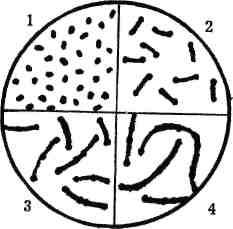
Коки (коккоз - зерно, кісточка) - кулясті мікроорганізми сферичної, еліпсоподібної, ланцетоподібної або бобовоподібної форми. За картиною розташування мікробів у мазках, яка залежить від способу поділу клітин і наступного їх розходження, кокоподібні бактерії поділяють на ряд груп (рис. 5).
Рис. 5. Основні форми бактерій:
1-6 сферичної форми: 1 - стафілококи; 2-3 диплококи; 4 - стрептококи; 5 - тет-ракоки; 6 - сарцини; 7-9 паличкоподібні; 10-12 спіралеподібні форми: 10 - вібріони; 11 - спірили; 12 - спірохети
Мікрококи (тісгоз - дрібний) характеризуються поодиноким і безладним розташуванням клітин. Як правило, це сапрофітні мікроорганізми і за звичайних умов не викликають захворювань у людини. Проте в осіб з імуноде-фіцитними станами або тих, хто переніс складні операції, трансплантації органів і тканин, вони можуть спричиняти бактеріемію, гнійно-септичні ускладнення.
Диплококи (сііріооз - подвійний) - група коків, які після поділу не розходяться, а існують парами. Типовими представниками є збудники епідемічного цереброспінального менінгіту та гонореї. Ці мікроорганізми мають характерну бобовоподібну форму і в мазках увігнутими сторонами повернуті один до одного. Збудники крупозної пневмонії та деяких інших гнійно-септичних процесів також належать до диплококів, але мають ланцетоподібну форму або форму полум'я свічки.
Стрептококи (зіхеріоз - намисто) - мікроби, які після поділу в одній площині не розходяться, а формують ланцюжки, що складаються з 3-4, а деколи й десятків клітин круглої або еліпсоподібної форми. Частина їх є сапрофітами, представниками нормальної
35
мікрофлори людини, інші викликають важкі гнійно-септичні процеси (пневмонію, менінгіт, остеомієліт, холецистит, сепсис тощо). Доказана роль стрептококів у розвитку скарлатини, ревматизму, бешихи.
Тетракоки (їеїга - чотири) - коки, які після поділу у двох взаємно перпендикулярних площинах утворюють тетради. Як правило, ці мікроорганізми непатогенні для людини.
Сардини (загсіпа - пака, тюк) - коки, у яких поділ відбувається в трьох взаємно перпендикулярних площинах, а після поділу вони не розходяться і розташовуються у вигляді паків з 8, 16, 32, 64 клітин. Патогенних представників серед них немає.
Стафілококи (зїарпуіе - гроно) - коки, які діляться в декількох площинах, клітини розташовуються хаотично, у вигляді скупчень, що в мазках із чистої культури нагадують виноградне гроно. Стафілококи - убіквітарні мікроби, зустрічаються в повітрі, грунті, організмі людини і тварин. Чисельні їх представники спричиняють різноманітні гнійно-септичні захворювання: фурункул, карбункул, гідроаденіт, флегмону, нефрит, холецистит, менінгіт, пневмонію, сепсис тощо. Практично в організмі людини немає органів і тканин, які не ушкоджуються стафілококами.
Не менш дивовижний світ паличкоподібних або циліндричних бактерій, які за своєю кількістю перевищують кокові мікроорганізми. Середні розміри їх 0,5-1,5 мкм завдовжки і 0,5-2 мкм завширшки.
Паличкоподібні бактерії мають різноманітну форму (циліндричну, еліпсоподібну, овальну, веретеноподібну, у вигляді барабанної палички або тенісної ракетки). їх кінці можуть бути рівні або нібито обрублені й навіть увігнуті (збудник сибірки), заокруглені (кишкові палички, збудники черевного тифу, дизентерії). Зустрічаються паличкоподібні форми із загостреними кінцями (фузобактерії), булаво-подібними потовщеннями на них. Часто трапляються мікроби, що мають розгалуження (мікобактерії туберкульозу). Це дозволяє розпізнавати вид мікроорганізмів, що має велике значення при лабораторній діагностиці.
Паличкоподібні бактерії, залежно від здатності утворювати спори та їх діаметру, поділяють на власне бактерії, бацили і клостридії.
Власне бактерії - мікрооганізми, які не здатні до утворення спор. До них належать збудники сальмонельозів, черевного тифу, дифтерії, туберкульозу, кашлюка.
Бацили (ЬасШиз - паличка) і клостридії (сіозїег - веретено) здатні утворювати спори. До першої групи належать збудники сибірки, до другої - збудники газової анаеробної інфекції, правця, ботулізму.
За аналогією з коками, залежно від способу розташування у мазках, паличкоподібні мікрооганізми поділяють на: а) монобактєрії (кишкова, черевнотифозна, дизентерійна палички, збудник чуми) і монобацили (збудники правця, ботулізму); б) диплобактерії (клебсієли пневмонії) і диплобацили; в) стрептобактерії (збудники м'якого шанкру) і стрептобацили (збудники сибірки).
Спіралеподібні бактерії мають звивисту, штопороподібну форму. До цієї групи бактерій належать вібріони, спірили, спірохети (рис. 5).
Вібріони ^іЬгаге - коливатись, тремтіти) - бактерії, з одним невеликим вигином розміром 1/4 завитка спіралі, що надає їм схожість із комою. Представниками цієї групи є збудники холери і холероподібні вібріони, які населяють водоймища.
Спірили (зріга - завиток, спіраль) - бактерії, які мають декілька вигинів, що надає їм форму штопора. Патогенним представником є спірила, яка викликає у людини содоку (хворобу укусу щурів). До цієї групи мікроорганізмів належать також кампілобактерії та гелікобактерії, які здатні спричиняти у людини захворювання шлунково-кишкового тракту, сечостатеве'! системи.
Спірохети (зреіга - виток, паііе - довге волосся) також мають штопороподібну форму і розрізняються між собою числом завитків і довжиною (довжина може сягати 500 мкм, а діаметр збудників -0,3-1,5 мкм). Серед них виділяють патогенні для людини трепонеми (їгеро - повертати, пето - нитка), борелії (за прізвищем французького бактеріолога А. Боррела) і лептоспіри (Іерїоз - тонкий, ніжний). Трепонеми викликають у людини сифіліс, лептоспіри - лептоспіроз, борелії - поворотний тиф. Існує велика група непатогених спірохет (власне спірохети, сапроспіри, кристиспіри).
Ниткоподібні бактерії для людини непатогенні. Тіобактерії та залізобактерії є мешканцями грунтів, водоймищ, беруть участь у процесах кругообігу речовин у природі. До цієї групи мікроорганізмів можна віднести й актиноміцети, які здатні викликати у людини тяжкі захворювання - актиномікози.
Описані морфологічні форми бактерій не обмежують існування мікробного царства. Деякі його представники можуть нагадувати кільце, шестикутну зірку, набувати червоподібної форми. Це, в основному, вільноживучі мікроорганізми, які беруть участь у процесах біо-деградації різноманітних природних сполук. Патогених представників серед них немає, і в курсі медичної мікробіології вони не розглядаються.
36
37
Ультраструктура бактеріальної клітини. Бактеріальні клітини є прокаріотичними живими системами. Між ними та еукаріотами (еи -справжній, кагуоп - ядро) існують суттєві відмінності, які дозволяють віднести представників мікробного світу до єдиного царства. Слід пам'ятати, що в еукаріотів тканини та органи складаються з окремих клітин, що знаходяться у фізіологічній метаболічній залежності і не можуть існувати окремо. Мікробна клітина - абсолютно автономний складний організм, здатний до самостійного, індивідуальнго існування. Найсуттєвішою ознакою прокаріотів є відсутність ядра. Його роль відіграє нуклеоїд - ядерна речовина, яка дифузно розташована в цитоплазмі та не відмежована від неї каріолемою. Нуклеоїд клітини складається з однієї нитки ДНК, замкненої в кільце, гістоноподібні
білки та ядерце відсутні. У бактерій немає таких органел, як міто-хондрії, апарат Гольджі, ендоплазматичний ретикулюм, хлоропласти, мікротільця. Проте вони мають мезосоми, функція яких аналогічна мітохондріальній. Константа седиментації мікробних рибосом складає 705, в той час як в еукаріотів -808. Існують також суттєві відмінності за будовою джгутиків, наявністю вакуолей тощо.
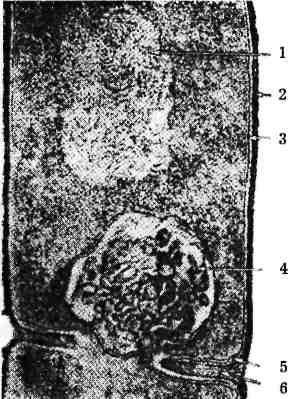
Рис. б. Ультратонкий зріз бактерії:
1 - нуклеоїд; 2 і 5 - клітинна стінка;
- цитоплазматична мембрана;
- мезосома; 6 - клітинна перегородка
38
Незважаючи на такі кардинальні відмінності в структурі клітин різних систем, загальний план їх будови залишається подібним. Прока-ріотний організм містить у собі майже всі клітинні елементи: оболонку, цитоплазму, ядерний апарат, включення (рис. 6). Нуклеоїд. Ядерний апарат бактеріальної клітини займає її центральну частину, має неправильну форму і не відмежовується від цитоплазми оболонкою, поєднується з цитоплазматичною мембраною і мезосомою. Він складається з однієї суперспіралізованої подвійної нитки ДНК діаметром до 2 нм, замкнутої в кільце, інтегрованої з РНК, РНК-полімеразою та білком у співвідношенні 1:1:3. Довжина цієї гігантської молекули може сягати до 1,5-3 мм. Молекулярна
маса нуклеоїда - (1-3)х10в дальтон, і містить він до 8x10" пар нуклеїнових основ. Вміст пар основ А+Т і Г+Ц в молекулі кожної клітини є постійним для певного виду бактерій, а частка Г+Ц у загальній молекулярній масі становить 23-75 %.
Як правило, в клітині нуклеоїд представлено однією копією, проте під час поділу клітини число цих копій може збільшуватись до 2-9.
Досить часто бактерії поруч із хромосомною містять позахромо-сомну ДНК значно менших розмірів, також скручену в кільце і локалізовану в цитоплазмі. Такі елементи одержали назву плазміди. Вони детермінують синтез деяких речовин, ферментів, забезпечують стійкість бактерій до антибіотиків, отже, надають їм певних селективних переваг.
Ядерну субстанцію мікробів можна виявити в ультратонких зрізах при дослідженні їх в електронному мікроскопі, за допомогою імунофлуоресцентної, радіоімунної мікроскопії, радіоавтографії, а також забарвлюючи її за методами Робіноу-Фельгена, Пікарського тощо.
Цитоплазма бактерійних клітин має рідку консистенцію, прозора, гомогенна, відмежовується від зовнішнього середовища цитоплазматичною мембраною. Вона є своєрідною колоїдною системою, що складається з різноманітних молекул білків, ліпідів, води, ДНК і РНК, вуглеводів, полісахаридів та інших сполук. В'язкість її у 800-8000 разів перевищує аналогічний показник води. Будова і консистенція цитоплазми залежить від віку мікроба - гомогенна у молодих клітин вона поступово перетворюється на дрібнозернисту структуру в старих, набуваючи вигляду щільників. У ній з'являються вакуолі, волокнисті утворення, збільшується її густина, за консистенцією вона ' нагадує гель.
При ультрацентрифугуванні цитоплазми можна одержати "розчинну" фракцію, до якої входять різноманітні ферменти, і фракцію "часток" з мембран та рибосом. Рибосоми виконують роль фабрики синтезу білка, їх розмір досягає 16x18 нм. Складаються вони з двох білкових субодиниць 308-508. Клітина може містити до 5000-50000 рибосом, число їх збільшується при активному синтезі білка. Часом рибосоми збираються у скупчення, які називають полірибосомами або полісомами.
Відмінності між рибосомами еукаріотичних та прокаріотичних організмів мають вирішальне значення у пошуках шляхів боротьби
39
■■'■
з останніми як збудниками інфекційних захворювань. Відомо, що деякі антибіотики здатні частково або повністю припиняти синтез білка саме на 705 рибосомах, залишаючи інтактними 803 рибосоми. У процесі життєдіяльності мікрооганізмів у цитоплазмі з'являються морфологічно диференційовані частки, які називають включеннями. Вони бувають різними за своєю природою і виконують різноманітні функції.
Запасні речовини прокаріотів представлено полісахаридами, ліпідами, поліпептидами, поліфосфатами, сіркою. Як полісахариди відкладаються крохмаль, глікоген, гранульоза. У несприятливих умовах вони забезпечують клітину вуглецем та енергією.
Ліпіди можуть накопичуватись у вигляді гранул Р-оксимасляної кислоти, їх можна побачити навіть при звичайній мікроскопії, забарвлюючи препарати Суданом III або Суданом чорним.
Широко розповсюджений тип поживних речовин - поліфосфати. Вони містяться у гранулах, які називають волютиновими, і використовуються клітинами як джерело фосфору. Крім того, вони мають макроергічні фосфатні зв'язки, отже, забезпечують потреби клітини в енергії. Зерна волютина називають ще метахроматичними включеннями, тому що вони забарвлюються в колір, невластивий основному барвнику. Наприклад, метиленова синька забарвлює їх у темно-фіолетовий колір, в той час як цитоплазму клітини - в голубий. Вперше включення такого типу було знайдено у ЗрігШит уоіиіапз, звідки вони й одержали таку назву. Наявність зерен волютину характерна для коринебактерій і, зокрема, для збудника дифтерії.
Аналогічну функцію забезпечення енергетичних потреб клітини можуть виконувати включення колоїдної сірки, а в деяких анаеробних мікроорганізмів вона виступає ще й донором електронів у біохімічних процесах.
Деколи у бактерій з'являються особливі утворення, які називають вакуолями. їм відводять роль сховища різноманітних бактеріальних ферментів, а також резервуара, де скупчуються непотрібні клітині продукти обміну.
Оболонка бактерій складається з цитоплазматичної мембрани, клітинної стінки і капсули.
Вміст клітини відмежовується від навколишнього середовища за допомогою цитоплазматичної мембрани (ЦПМ) - м'якого, пластичного утворення. її будова у прокаріотів і еукаріотів подібна, що свідчить про існування універсальної "елементарної мембрани". Відрізняються
40
вони тільки відсутністю стеролів у перших. Мембрана - обов'язковий структурний компонент мікробної клітини, без неї вони гинуть. За хімічним складом вона є білково-ліпідним комплексом із невеликою кількістю вуглеводів. Формуючи всього 8-15 % маси клітини, мембрана містить до 70-90 % її ліпідних субстанцій.
Дослідження під електронним мікроскопом показали, що мембрана є багатошаровим утворенням. Вона складається з подвійного шару фосфоліпідних молекул. Гідрофобні їх кінці (фосфоліпіди та тригліце-риди) спрямовані всередину, а гідрофільні "головки" - назовні. Такий тип розташування стабілізує мембрану. В цей шар вмонтовано інтегральні білки, які пронизують його наскрізь. Деякі групи білків прикріплюються до поверхні мембрани, тому їх називають периферійними. Деколи мембрана покривається ще одним особливим типом білка - поверхневим.
Елементарна мембрана здатна утворювати інвагінати, які називаються мезосомами. Вони відіграють велику роль у життєдіяльності клітини, постачаючи її енергією та беручи активну участь у процесах реплікації нуклеоїда.
Функції мембранного комплексу різноманітні: він забезпечує селективну проникність та транспорт різноманітних речовин іззовні всередину і навпаки завдяки існуванню в ньому особливих білків-ферментів пермеаз; здійснює транспорт електронів та окисне фос-форилювання субстратів; генерує електрохімічну енергію трансмембранного потенціалу; виділяє гідролітичні ферменти; проявляє біосинтетичну активність; є місцем прикріплення джгутиків.
Виявити цитоплазматичну мембрану можна в ультратонких зрізах бактерій під електронним мікроскопом.
Деякі поверхнево-активні речовини і антибіотики (поліміксини) здатні руйнувати мембрану і викликати загибель клітини. Це використовується в пошуках оптимальних шляхів боротьби зі збудниками інфекційних хвороб.
Клітинна стінка. Клітинна стінка створює захисний шар, який врівноважує високий внутрішній осмотичний тиск бактерій (5-20 атмосфер). Таку міцність забезпечує речовина - муреїн, пептидоглікан. Він складається з особливих полімерних ланцюгів, у яких чергуються залишки ІЧ-ацетилмурамової кислоти і М-ацетилглюкозаміну, в свою чергу сполучених між собою Р-1,4-глікозидними зв'язками. Залишки мурамової кислоти з'єднуються пептидними зв'язками із тетра-иептидами амінокислот: Ь- і Б-аланіну, Б-глутамінової та
41
мезодіамінопімелінової кислот, Ь-лізину. Пептидними містками такі гетерополімерні ланцюги зв'язуються між собою, утворюючи гігантський муреїновий мішок. Те, що до складу бактерій входять речовини, відсутні в тваринних і рослинних клітинах (гТ-ацетил-мурамова кислота і М-ацетилглюкозамін), створює можливість цілеспрямованого знищення бактерій деякими антибіотиками (пеніциліни, цефалоспорини), оскільки клітинні стінки еукаріотів при цьому не пошкоджуються.
Створена з муреїну структура виконує функцію опорного каркасу, надаючи форму мікробній клітині, крім того, з ним зв'язуються інші речовини.
За особливостями будови мікробного мурешового каркасу і вмістом деяких речовин у клітинній стінці можна відрізнити так звані-грампозитивні бактерії від грамнсгативних. Поділ їх на ці дві групи було запропоновано у 1884 р. Христіаном Грамом, який звернув увагу на особливості фарбування мікробів.
У грампозитивних бактерій муреїновий шар складає 30-70 % маси клітинної стінки, утворюючи до 40 шарів. Замість мезодіамінопімелінової кислоти в ньому міститься ЬЬ-діамінопімелінова кислота або лізин. Суттєвою особливістю є наявність особливих тейхоєвих кислот.
Під електронним мікроскопом таку клітинну стінку видно як губчасту структуру з порами діаметром 1-6 нм.
Грамнегативні бактеріальні клітини мають значно складнішу будову стінки. До її складу входить більше розмаїття біологічних молекул. Муреїновий шар у них одношаровий, складає до 10 % маси сітки. Він містить мезодіамінопімелінову кислоту, немає лізину, а міжпеп-тидні містки відсутні. Тейхоєвих кислот у стінці також немає. Зовні до муреїнового шару прилягає шар ліпопротеїну, який переходить у зовнішню мембрану, що складається з білків, фосфоліпідів і ліпополісахаридів, типових для елементарних мембран. Над мембраною, інтегруючись із нею, розміщується ліпополісахарид. Він має внутрішнє й зовнішнє полісахаридне ядро, пов'язане з ліпідом А. За зовнішніми специфічними боковими ланцюгами ліпополісахаридів збудники можна диференціювати один від іншого, що використовується при ідентифікації. Ліпід А забезпечує токсичні властивості мікробної клітини, викликаючи в людини підвищення температури, пронос та інші прояви хвороби.
Клітинна стінка крім опорної та захисної виконує ще ряд важливих функцій. Зокрема, вмонтовані у фосфоліпідний шар трансмембранні
42
білки (порини) - це заповнені водою канали, через які проходять низькомолекулярні сполуки. Периплазматичний простір між цитоплазматичною мембраною та клітинною стінкою у грамнегативних бактерій виступає сховищем для різноманітних ферментів - деполі-мераз, протеїназ, нуклеаз, рестрикційних ферментів, відіграє роль у забезпеченні осморегуляції.
Під впливом різноманітних речовин клітинна стінка може бути знищена. Так, при дії лізоциму на суспензії грампозитивних мікрококів виникає їх швидкий лізис і просвітління середовища. Аналогічний ефект спричиняє пеніцилін. Лізоцим розриває в муреїні глікозидні зв'язки, а пеніцилін попереджує утворення пептидоглікану, що супроводжується руйнуванням клітинної стінки. При цьому утворюються чутливі до осмотичних умов округлі клітини - протопласти, у яких повністю втрачена клітинна стінка. При дії вказаних препаратів на грамнегативні бактерії формуються клітини, які зберігають рештки клітинної стінки. їх називають сферопластами.
Протопласти і сферопласти належать до субклітинних форм бактерій. Вони мають круглу форму, високочутливі до осмотичних умов середовища, утворюють спори, якщо був ініційований процес споруляції, але не мають здатності до розмноження. Сферопласти, на відміну, від протопластів, здатні адсорбувати на своїй поверхні бактеріофаги і відновлюватись у вихідні форми при відміні дії чинників, які викликали їх утворення.
В організмі людини і тварин при антибіотикотерапії (пеніцилін, бацитрацин, новобіоцин) створюються умови для порушення синтезу пептидоглікану, а бактерійні клітини, втрачаючи свою клітинну стінку, перетворюються в Ь-форми. Це клітини до 50 мкм у діаметрі, які зберігають тенденцію до перетворення у вихідні форми, продукують токсини, гіалуронідазу. Описані такі форми бактерій у збудників гонореї, туберкульозу, черевного тифу, бруцельозу та інших. Вони здатні викликати у людини захворювання, які супроводжуються тривалим перебігом. Утворення таких форм мікроорганізмів вважають за спосіб переживання несприятливих факторів зовнішнього середовища.
Слід відмітити, що в природі існують мікроорганізми, у яких немає клітинної стінки. Вони називаються мікоплазмами і мають виражені патогенні властивості, викликаючи різноманітні захворювання дихальної, серцево-судинної, сечо-видільної систем.
Клітинну стінку мікробів можна зруйнувати лугом, ультразвуком, механічним методом. Виявити її можна різними способами:
43
спостерігаючи в електронному, фазовоконтрастному та аноптрально-му мікроскопах, при мікроскопії автолізованих бактерій, застосовуючи спеціальні методи забарвлення (шафраніном, синьою вікторією). У лабораторних умовах легко дослідити наявність клітинної стінки, використовуючи явище плазмолізу. При цьому клітину занурюють у гіпертонічний розчин хлориду натрію або 0,2М розчин нітрату калію. Вода за градієнтом концентрації виходить із клітини назовні, цитоплазматична мембрана разом із цитоплазмою зморщуються, відшаровуючись від стінки, яка зберігає форму бактеріальної клітини і стає видимою під мікроскопом.
Капсула. Зовні бактеріальна клітина може бути вкрита речовиною слизового характеру. Вона не має для мікроба життєзабезпечуючого значення, однак захищає його від дії несприятливих факторів зовнішнього середовища, надає стійкості до фагоцитозу, захищає від про-никненя бактеріофагів, забезпечує вірулентні властивості збудників (рис. 7). За своєю хімічною будовою капсула належить до полісаха-Рис 7. Капсули бактерій ридних субстанцій, а у В. апїЬгасіз - до білкових. Характерним для капсули є наявність великої кількості води. У деяких мікроорганізмів капсула утворюється тільки в організмі тварин або людини (пневмококи, збудники сибірки, чуми та ін.), у інших (клебсієли) вона присутня постійно, навіть коли збудник росте на живильному середовищі. Інколи капсула оточує разом декілька клітин (сибіркова бацила), тоді такі утворення називають зооглеями. Стафілококи, збудники дифтерії, деякі стрептококи та інші бактерії здатні утворювати мікрокапсули, особливо при культивуванні на середовищах, багатих на вуглеводи.
Капсулу можна розглядати у звичайному світловому мікроскопі, якщо забарвлювати нативні препарати простим методом. Однак для виявлення капсул частіше використовують метод Буррі-Гінса, при якому фон препарата створюють тушшю, а мікроорганізм додатково забарвлюється фуксином. У таких випадках на темному фоні видно червону паличку, яка оточена світлим ободком - капсулою. Джгутики. Поверхня тіла мікроорганізмів може бути вкрита особливими виростами, що називаються джгутиками, які забезпечують локомоторну функцію. їх число, спосіб розміщення, довжина є постійними ознаками для певного виду бактерій, що враховується
44
при проведенні систематики прокаріо-тичних організмів (рис. 8, 9).
Довжина джгутиків досягає 20 мкм, тоді як товщина - всього 12-18 нм, що лежить за межами роздільної здатності мікроскопа. Джгутики бактерій складаються із спірально закручених ниток особливого білка флагеліну, який утворює спіраль навколо внутрішнього порожнистого простору. У них виділяють три основні частини: спіральну нитку, гак та ^ 8 Джгутики бактерій:
базальне ТІЛО (два - ЧОТИри спеціальних 1 - монотрихи; 2 - амфітрихи; КІЛЬЦЯ 3 Центральним стержнем), За 3 - лофотрихщ 4 - перитршси
допомогою яких джгутик закріплюється у цитоплазматичній мембрані та клітинній стінці).
Джгутики асоціюють з білковим Н-антигеном мікробної клітини, визначення якого має певне значення в лабораторній діагностиці інфекційних хвороб.
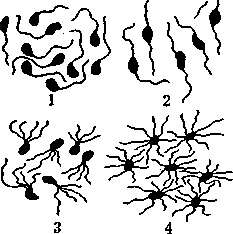
За способом розташування джгутиків мікроорганізми поділяються на ряд груп. Монотрихи - бактерії, що містять джгутик на одному з полюсів клітини (холерний вібріон). Такі мікроорганізми найрухоміші серед інших: за 1 с вони здатні переміщуватись на віддаль, яка у 20 разів перевищує довжину їх тіла. Лофотрихи мають пучок джгутиків на одному з полюсів (псевдомонади, Васіїїиз тедаїегіит). У амфітрихів джгутики або їх пучки розташовані на обох полюсах (спірили). Пе-ритрихи мають джгутики, розміщені по всій поверхні тіла мікроба (протей, ешерихії, сальмонели), число їх може досягати 1000.
Виявити джгутики можна за допомогою прямих та непрямих методів. При непрямих методах спостерігають за рухом мікроорганізмів у темному полі мікроскопа, у "висячій" чи "роздавленій" краплях, за допомогою фазово-контрастної, аноптральної мікроскопії. За допомогою прямих методів джгутики забарвлюють барвниками, або солями металів, попередньо наносячи на них протраву для збільшення у розмірах, або досліджують на ультратонких зрізах під електронним мікроскопом.
Рух джгутиків забезпечується енергією трансмембранного потенціалу, яка генерується на цитоплазматичній мембрані. У більшості мікробів з полярним розташуванням джгутиків вони обертаються зі швидкістю 3000 обертів за 1 хв.
45
Джгутики
мають пристосувальне значення. Вони
надають клітині змогу переміщуватись
у рідкому середовищі в пошуках більш
сприятливих умов існування. У відповідь
на зовнішні подразнення (хімічні
речовини, температура тощо) або спонтанно
здатні змінювати характер свого
обертання і напрям руху. Це називають
таксисом. Відповідно до факторів, що
його викликають, розрізняють хемотаксис,
фототаксис, аеротаксис.
До поверхневих структур бактерійної клітини належать різноманітні придатки, які позначають терміном пілі (рііиз - волос): стебельця, шипики тощо. Особливим типом таких утворень є ворсинки, фімбрії (іітЬгіа - торочка). їх число може перевищувати 10000, довжина сягає до 2 мкм, а товщина -3-25 нм. Як і джгутики, вони складаються з білкових субодиниць. Описані ворсинки двох класів. Загальні ворсинки (1-го класу) забезпечують ад-Рис. 9. Джгутики й гезію (прикріплення) мікроба до субстрату, через
ворсинки бактерій НИХ Всередину КЛІТИНИ МОЖуТЬ Проникати ДЄЯКІ
метаболіти і навіть бактеріофаги. Ворсинки 2-го класу (статеві) беруть участь у передачі генетичної інформації від клітини до клітини при кон'югації (рис. 9).
Спороутворення. На певній стадії свого розвитку, коли запаси поживних речовин вичерпуються, бактерії всередині формують спору (ендоспору) округлої форми. Від вегетативних форм вони відрізняються пригніченням функціонування генетичного апарата, майже повною відсутністю обміну речовин, малою кількістю вільної води, підвищеною концентрацією іонів кальцію, появою у складі дипіколіно-вої кислоти, з якою пов'язують термостійкість спор. Для них характерна поява додаткових оболонок, які запобігають дифузії і проникненню речовин іззовні, більш висока стійкість до пошкоджуючих факторів зовнішнього середовища і здатність тривалий час зберігати свою життєздатність. Спори утворюють два роди грампозитивних паличок - Васіїїив (спора за діаметром менше поперечника палички) і Сіозігігіішп (спора перевищує розміри палички) і один рід грампозитивних коків (Зрогозагсіпа).
Спори утворюються тільки в зовнішньому середовищі, в організмі тварин та людини процесу споруляції не відбувається. Вони мають еволюційне значення, забезпечуючи збереження виду, а не виконуючи функцію розмноження (рис. 10).
46
С пороутворення
починається, коли в оточуючому клітину
середовищі зникають джерела азоту та
вуглецю.
пороутворення
починається, коли в оточуючому клітину
середовищі зникають джерела азоту та
вуглецю.
Спочатку в клітині при інвагінації мембрани виділяється особливе термінальне ядро, яке містить один клітинний геном, компоненти апарата синтезу білка і власну енергетичну систему. ВОНО ВКриваЄТЬСЯ Власною мембраною та Риа 10- Розташування
мембраною материнської клітини, які утворю- П^Г^Їб-ють стінку спори. Вона складається з нормаль- термінальне ного пептидоглікану. Стінку оточує кора, яка містить незвичний пептидоглікан з малим числом поперекових з'єднань і чутливий до лізоциму (автоліз його відіграє вирішальну роль у процесі проростання спори). Оболонка спори складається з кератиноподібного білка і зумовлює погану проникність і стійкість його до хімічних речовин. Екзоспорій оточує всю спору і складається з ліпопротеїнів.
Індукція спороутворення відбувається протягом декількох годин. Розрізняють декілька стадій: підготовчу, передспори, утворення оболонок і дозрівання.
Процес проростання спори також відбувається у декілька етапів. Під час фази активації діючий фактор пошкоджує оболонку спори (висока температура, механічні пошкодження, зміна рН середовища в кислу сторону). На ініціальній (початковій) стадії спора за сприятливих умов починає проростати, поглинає воду, руйнується пептидоглікан кори. На стадії росту внаслідок руйнування оболонок утворюється нова вегетативна клітина. Процес триває протягом 4-6 годин.
Залежно від локалізації спор виділяють такі типи їх розміщення: центральне (сибіркова бацила), субтермінальне, коли спора розташовується ближче до одного з кінців, тоді паличка нагадує тенісну ракетку (збудники ботулізму), і термінальне, коли спора знаходиться на кінці мікроба, надаючи йому вигляду барабанної палички (збудники правця).
Спори тривалий час зберігаються в стані анабіозу. У грунті вони южуть перебувати понад 200 років. Знайдено спорові мікроор-
ізми у льодовиках Антарктиди, вік яких оцінюється в 12 тис юків.
Ендоспори бактерій характеризуються високою термостійкістю. априклад, .спори сінної палички витримують кип'ятіння протягом год. Саме резистентність спор примусила мікробіологів винайти
47
автоклав, розробити надійні способи стерилізації. При автоклавуванні під тиском 1,5-2 атмосфери і температурі 127-132 °С спори гинуть.
Наявність спор у бактерій може мати діагностичне значення, а також спричиняє вибір тактики при знезаражуванні хірургічного інструментарію та перев'язного матеріалу.
Спори мають здатність сильно заломлювати світло, тому на незафарбованих препаратах їх видно у вигляді блискучих зерен. У зв'язку з тим, що вони стійкі до дії несприятливих факторів зовнішнього середовища, забарвлювати їх досить важко. Найчастіше використовується метод Ожешко, при якому спори попередньо протравлюють соляною кислотою, а потім забарвлюють за методом Ціля-Нільсена. Спори при цьому набувають червоного, а вегетативна клітина - голубого забарвлення.
Морфологічні особливості окремих груп мікроорганізмів
Актиноміцети. Актиноміцети (акііз - промінь, тукез - гриб) -одноклітинні бактерії, які раніше вважали грибами через здатність клітин галузитись. Вони належать до родин Асйпотусеїасеае та Мосагйіасеае, до складу яких входить понад 400 різноманітних видів. Будова актиноміцетів подібна до структури звичайної мікробної клітини, однак існують деякі відмінності за хімічним складом цукрів. Актиноміцети, як правило, сапрофіти, вільно живуть у грунті, забезпечуючи його плодючість і запах, та інших об'єктах зовнішнього середовища. Багато з них є продуцентами антибіотиків (стрептоміцин, тетрацикліни).
Проникаючи в організм людини або тварин, за певних умов актиноміцети здатні викликати захворювання з хронічним перебігом -актиномікоз та нокардіоз. Збудниками їх є Асііпотусез ізгаеііі, А.Ьоуіз, Апаезіипйіі, Мосагсііа азіегоісіез. Найчастіше ушкоджуються шкіра, підшкірна клітковина з утворенням нориць, лімфатичні вузли, органи дихання. У тканинах, уражених актиноміцетами, знаходять особливі структури - скупчення видозміненого міцелію, які називають друзами.
Патогенні спірохети. Це особлива група грамнегативних звивистих мікроорганізмів, які мають вигляд довгих, спірально закручених, тонких ниток. Довжина їх коливається в межах 7-50, а
товщина - 0,3-0,5 мкм. Вони належать до порядку Зрігоспаеіаіез, який об'єднує три патогенних для людини роди: Тгеропета, Вогге-Ііа і Ьеріозріга, які відрізняються між собою за рядом ознак.
Спільним для всіх спірохет є наявність цитоплазматичного циліндра, відмежованого ЦПМ і вкритого тришаровою клітинною стінкою. Він намотаний навколо фібрилярного апарата, що складається з білка флагеліну, і розташований всередині клітини. Фібрили прикріплюються до дисковидних утворень на кінцях збудників, які називаються блефаропластами. У цитоплазмі знаходиться нуклеоїд, рибосоми, мезосоми, включення. Спор і капсул мікроби не утворюють, однак при несприятливих умовах здатні формувати цисти. Спірохети мають різні типи рухів: згинальний, поступальний, обертальний, маятникоподібний.
Число і форма завитків характерні для ~ кожного виду спірохет.Вони формують завитки , І порядку. У трепонем є 8-14 завитків І порядку, однакових за формою та величиною. борелій нараховується 4-8 великих завитків « ,
неправильної форми, неоднакових за величи-
ною. Ці мікроорганізми найбільші серед патогенних спірохет. Лептоспіри мають 12-18 завитків І порядку, але вони дрібні й тісно прилягають один до одного, під звичайним світЛОВИМ МІКРОСКОПОМ ПраКТИЧНО НЄ ДИфереНЦІЮються. На кінцях мікроби мають вторинні завитки, що надає їм 5-, С-подібної форми, робить схожими на гачок
За методом Романовського-Гімзи лептоспіри забарвлюються в блідо-рожевий колір, трепонеми - в рожевий, борелії - в синьо-фіолетовий.
Патогенними для людини є Тгеропета раїїійит, яка викликає сифіліс, Воггеїіа гесиггетлз - збудник епідемічного поворотного тифу, В. сїииопі, В. саисазіса - збудники ендемічного поворотного тифу, а також Ьерїозріга іпіеггодапз, що спричиняє у людини лептоспіроз.
Рикетсії. Це прокаріотичні мікроорганізми з родини Кіскеизіа-сеае, що одержали свою назву від прізвища американського дослідника Г. Рікетса, який загинув від лабораторного зараження висипним тифом.
48
49
Рикетсії - дрібні плеоморфні бактерії розміром 0,3-0,6 х 0,4-2 мкм (рис. 12). їх поділяють на чотири морфологічних типи: кокоподібні, паличкоподібні, бацилярні та ниткоподібні.
Вони мають типову структуру бактеріальної клітини. У них є оболонка, цитоплазма, нуклеоїд. Будова їх клітинної стінки ідентична грамнегативним бактеріям.
Рис. 12. Чотири морфологічні РИКЄТС* Нерухомі, не утворюють СПОр і
типи рикетсій: і - кокотюдіб- К£іПСУл- Вони містять велику кількість ліпі-ні; 2 - паличкоподібні; з - ба- Дів (до 46 %), тому використання методу цилярні; 4 - ниткоподібні Грама для їх фарбування обмежене. Найчастіше для цього слугують методи Романовського-Гімзи (мікроби фарбуються в голубий колір, а ядерна субстанція - в червоний), Здродовського (рикетсії рубіново-червоні, а клітинні елементи -блідо-голубі), негативне контрастування Айзенберга (незафарбовані рикетсії чітко виявляються на голубому фоні).
На поживних середовищах рикетсії не ростуть, їх можна культивувати в організмі лабораторних тварин, у жовтковому мішку курячого зародка, у легенях білих мишей, у кишечнику вошей.
Рикетсії - унікальні прокаріотичні мікроорганізми. У них поєднуються біологічні властивості прокаріотичних живих систем і вірусів. З першими їх єднає типова будова клітини, а з вірусами - те, що вони є облігатними внутрішньоклітинними паразитами, а також високий вміст ліпідів у цитоплазмі та клітинній стінці.
Більшість рикетсій непатогенні для людини. Вони паразитують на членистоногих. Однак деякі види здатні викликати важкі захворювання у людей, які називаються рикетсіозами. Серед них висипний тиф, марсельська лихоманка, гарячка Цуцугамуші, Ку-гарячка та інші. Збудниками цих та інших рикетсіозів є КіскеШіа ргоуагекіі, К. ІурЬі, К. їзшзиеатизпі, Сохіеііа ЬигпеШ.
Велику роль у передачі та розповсюдженні рикетсіозів відіграють воші, блохи та кліщі.
Хламідії. Хламідії є дрібними, кокоподібними, нерухомими бактеріями. Вони аспорогенні, не утворюють капсул, грамнегативні. Належать до родини СЬІатусІіасеае. Не ростуть на живильних середовищах, є облігатними внутрішньоклітинними паразитами. Поза клітиною людини вони існують у вигляді маленьких тілець діаметром
50
до 0,3 мкм. Протягом свого життєвого циклу хламідії проходять три стадії існування в клітині, утворюючи елементарні, ініціальні та проміжні тільця. Тривалість циклу розвитку хламідії - 36-72 год.
Хламідії (Спіатусііа Іхаспотаіїз, С. рзіМасі) здатні викликати небезпечні захворювання у людини: трахому, паховий лімфаграну-льоматоз, уретрити, кон'юнктивіти новонароджених, бленорею з включеннями, орнітоз, пневмонію, поліартрит, гастроентерит, ме-нінгоенцефаліт.
Мікоплазми. Мікоплазми є аспорогенними прокаріотичними організмами, які не мають ригідної клітинної стінки. Вони - поліморфні, сферичні або овоїдні утворення діаметром до 0,2 мкм, однак бувають і клітини діаметром до 1,5 мкм і більше. Мікоплазми не утворюють спор, грамнегативні. Вони мають типову будову бактеріальної клітини, але в них відсутня клітинна стінка.
Мікоплазми вибагливі до живильних середовищ, вимагаючи додавання до них холестеролу, нативного білка, пуринових та піримі-динових основ та інших компонентів.
Вони часто зустрічаються в стічних водах, грунті, паразитують в організмі людини і тварин. У людини вони спичиняють пневмонії, бронхіоліти, ангіни, уретрити, простатити, ураження придатків матки, артрити, ендокардити тощо. Найчастіше захворювання викликають Мусоріазта рпеитопіае, М. потіпіз, Цгеаріазта игеаіуіісит.
Патогенні найпростіші. Об'єктом дослідження в мікробіології виступає група еукаріотичних мікроорганізмів, які називаються найпростішими. Вони належать до типу Ргоїогоа і, відповідно, класів саркодових, джгутикових, споровиків та інфузорій.
Найпростіші - високоорганізовані живі системи з усіма функціями, притаманними тваринному організму розмірами від 2 до 150 мкм.
Будова їх складніша за бактерійну клітину. Вони мають чітко уособлене одне або декілька ядер з каріолемою І ядерцями, цитоплазму, яка відмежовується пеликулою, особливим видом еластичної мембрани. У них є ендоплазматичний ретикулюм, апарат Гольджі, мітохондрії, лізосоми, різні типи вакуолей як орган травлення. Деякі збудники мають особливі опорні фібрили. Рухаються вони за допомогою псевдоподій, джгутиків або війок.
Найпростіші мають складні цикли розвитку. Здатні розмножуватись безстатевим (поділом) і статевим шляхами або їх поєднанням При несприятливих умовах вони утворюють цисти, які здатні довго зберігатись, не втрачаючи патогенних властивостей.
51
Найпростіші широко розповсюджені у природі. Більшість із них непатогенні для людини. Однак деякі представники мають високо-інфекційні властивості та спричиняють амебіаз, трипаносомоз, лейшманіоз, урогенітальний та кишковий трихомоноз, токсоплазмоз, малярію, балантидіаз. Всі ці захворювання характеризуються тривалим і важким перебігом, ураженням численних органів і систем.
Патогенні гриби. Гриби - особлива велика група еукаріотів (понад 100000 видів), широко розповсюджена в природі, а грибкові захворювання (мікози) складають значну частину інфекційної патології людини.
Гриби представляють собою нефотосинтезуючі організми, які ростуть у вигляді ниток, що переплітаються та галузяться. їх унікальність полягає в тому, що вони подібні за деякими ознаками як до тваринних, так і до рослинних організмів. Із тваринним світом їх зближує наявність особливої полісахаридної субстанції - хітину, участь в обміні азоту сечовини, а в обміні вуглеводів - глікогену. Однак за способом живлення - шляхом всмоктування, а не заковтування їжі, а також необмеженим ростом, вони нагадують рослини.
Клітини більшості грибів вкриті твердою оболонкою, основу якої складають азотисті та безазотисті полісахарди та целюлоза. Цитоплазматична мембрана щільно прилягає до внутрішньої поверхні клітинної оболонки. В цитоплазмі міститься чітко диференційоване одне або декілька ядер з ядерцями, центральна вакуоль, мітохонд-рії, мікросоми, лізосоми, рибосоми, комплекс Гольджі, пластиди, секреторні гранули, різноманітні включення - волютин, солі органічних кислот, фосфатів, солей калію, натрію, заліза, глікоген, пігменти різнобарвних відтінків.
Молоді клітини грибів, як правило, яйцевидні, дещо видовжені, зрілі стають циліндричними, а старі - грушоподібними, булавоподіб-ними, веретеноподібними. Основу тіла гриба становлять особливі трубчасті нитки - гіфи, сукупність яких називають міцелієм. У гіфах нижчих грибів починають з'являтись перегородки, які є характерною ознакою у вищих грибів. Вони не суцільні, мають пори, через які здійснюється загальний обмін речовин. Кінцеві розгалуження міцелію мають своєрідну форму, за якою можна диференціювати окремі види. Так, гіфи міцелію дерматофітів можуть нагадувати роги оленів, канделябри, спіралі, булаву, гребінь півня тощо.
Розмноження грибів здійснюється безстатевим та статевим шляхом, що дозволило поділити їх на дві принципово відмінні групи -
недосконалі та досконалі гриби. Вегетативне розмноження здійснюється брунькуванням, кусочками грибниці, спорами, які виникають внаслідок розчленування гіфів міцелію. Спори можуть розміщуватись по боках або на кінцях міцелію (екзоспори), часто в особливих утвореннях, які називаються асками або сумками (ендоспори). Статеве розмноження здійснюється шляхом злиття чоловічих та жіночих гамет, внаслідок чого утворюється зигота, яка дає початок новому грибу.
Гриби - аеробні істоти. Для їх живлення необхідні численні азотисті, вуглецевмісні та мінеральні речовини. Вони мають різноманітні ферменти, що використовується для диференціації окремих видів.
Існують різні класифікації грибів, в основу яких покладено їх природну ознаку - будову органів плодоносіння, характер плодових тіл, їх форму, величину, морфологічні особливості.
Виділяють нижчі та вищі гриби. До нижчих належать хітридіомі-цети, ооміцети, зигоміцети. До вищих - аскоміцети, базидіоміцети та дейтероміцети.
Нижчі гриби непатогенні для людини. Однак окремі види мукорової плісняви рідко уражають шкіру, очі, зовнішні слухові проходи, легені, мозкові оболонки, шлунково-кишковий тракт, здатні викликати алергію, професійні оніхомікози та пароніхії у людей, які збирають і обробляють апельсини. Захворювання називаються зигомікозами.
Аскоміцети - клас сумчастих грибів, тому що спори їх зберігаються в особливих сумках - асках, об'єднує понад 300 різноманітних видів. Вони мають міцелій з вираженими перегородками. Патогенними для людини є представники родів Азрегдіїїиз, РепісШіиш, які викликають відповідно аспергільози та пеніциліози шкіри і внутрішніх органів зовнішніх слухових проходів, а також алергічні реакції.
Велике значення в медичній практиці мають гриби роду Сап-гіісіа. Вони часто є представниками нормальної мікрофлори організму людини. Але при важких хронічних захворюваннях, імунодефіцитах, злоякісних новоутвореннях, нераціональному застосуванні антибіотиків, гіповітамінозах та авітамінозах здатні викликати серйозні захворювання - кандидомікози. Молочниця у новонароджених дітей, заїди в осіб будь-якого віку, системні кандидози з ураженням шкіри, органів дихальної системи та шлунково-кишкового тракту із смертельними випадками - все це спектр дії одних і тих же дріжджеподібних грибів Сапсіісіа.
52
53


Великого
значення в медичній патології набула
група недосконалих грибів,
дейтероміцетів, до складу якої входить
багато видів. Вони мають септований
міцелій, їх життєвий цикл відбувається
в гаплоїдному стані, без зміни ядерних
фаз, статевого шляху розмноження у
них ще не описано.
Ця група грибів надзвичайно різноманітна і часто є причиною розвитку в людини важких захворювань - дерматофітій, глибоких мікозів. Серед них найчастіше зустрічаються епідермофітія стоп, мікроспорія, трихофітія, фавус.
Важкий і хронічний перебіг мають у людини такі мікози, як гістоплазмоз, криптококоз, північноамериканський бластомікоз або хвороба Джілкрайста-Стекса, пневмоцистоз у людей із синдромом набутого імунодефіциту та багато інших.
Однак гриби мають і велике корисне значення. їх використовують у харчовій, парфюмерній, хімічній промисловості як продуценти різноманітних ліполітичних та інших ферментів. Із представників родів РепісіІИшп, СерЬаІозрогіит одержують антибіотики (пеніциліни, цефалоспорини).
Матеріали до практичних занять