

Экоцитология.Лаб.-практ.9.Комплекс Гольджи
.docxЛабораторно-практическое занятие № 9
Тема: «Аппарат (комплекс) Гольджи»
Цель занятия: выявить морфо-функциональные особенности комплекса Гольджи.
Вопросы для обсуждения
1. Тонкое строение аппарата Гольджи.
2.
Демонстрационные препараты
Оборудование
1. Фотографии, схемы, рисунки из Атласа по биологии клетки, Ж.-К.Ролана, А. Селоши, Д. Селоши, пер.В.П. Белого, под ред. Ю.С. Ченцова. ─ М.: Мир. 1978. ─ 119 с.
Теоретическое обоснование для подготовки к занятию
Аппара́т (ко́мплекс) Го́льджи ─ мембранная структура эукариотической клетки, органелла, в основном предназначенная для выведения веществ, синтезированных в эндоплазматическом ретикулуме. Аппарат Гольджи был назван так в честь итальянского учёного Камилло Гольджи, впервые обнаружившего его в 1897 году (Fabene P.F., Bentivoglio M., 1998).

Рис. 1. Схема Аппарата Гольджи (А). Строение Аппарата Гольджи (Б)
Примечание: аппарат Гольджи ─ окруженные мембранами полости (цистерны) и связанная с ними система пузырьков. Функции ─ накопление органических веществ; «упаковка» органических веществ; выведение органических веществ; образование лизосом.
Аппарат (комплекс) Гольджи представляет собой стопку дискообразных мембранных мешочков (цистерн), несколько расширенных ближе к краям, и связанную с ними систему пузырьков Гольджи. В растительных клетках обнаруживается ряд отдельных стопок (диктиосомы), в животных клетках часто содержится одна большая или несколько соединенных трубками стопок.
В Комплексе Гольджи выделяют 3 отдела цистерн, окруженных мембранными пузырьками:
1. Цис-отдел (ближний к ядру).
2. Медиальный отдел.
3. Транс-отдел (самый отдаленный от ядра).
Эти отделы различаются между собой набором ферментов. В цис-отделе первую цистерну называют «цистерной спасения», так как с ее помощью рецепторы, поступающие из промежуточной эндоплазматической сети, возвращаются обратно. Фермент цис-отдела: фосфогликозидаза (присоединяет фосфат к углеводу ─ маннозе).
В медиальном отделе находится 2 фермента: манназидаза (отщепляет манназу) и N-ацетилглюкозаминтрансфераза (присоединяет определенные углеводы ─ гликозамины).
В транс-отделе ферменты: пептидаза (осуществляет протеолиз) и трансфераза (осуществляет переброс химических групп).
Тонкое строение аппарата Гольджи (АГ). В электронном микроскопе видно, что аппарат Гольджи представлен мембранными структурами, собранными вместе в небольшой зоне (рис. 1, 2); в виде стопки расположены плоские мембранные мешки (цистерны), количество таких мешков в стопке обычно не превышает 5-10. Между которыми располагаются тонкие прослойки гиалоплазмы. Каждая отдельная цистерна имеет диаметр около 1 мкм и переменную толщину; в центре мембраны могут быть сближены (25 нм), а на периферии иметь расширения ─ ампулы, ширина которых непостоянна.

Рис. 2. Схема строения диктиосомы (по Ченцову Ю.С., 2010)
Примечание: П ─ проксимальная (цис-) часть; Д ─ дистальная (транс-) часть; В ─ вакуоли; Ц ─ плоские мембранные цистерны; А ─ ампулярные расширения цистерн.
У некоторых одноклеточных их число может достигать 20 штук. Кроме плотно расположенных плоских цистерн в зоне АГ наблюдается множество вакуолей. Мелкие вакуоли встречаются главным образом в периферических участках зоны АГ; иногда видно, как они отшнуровываются от ампулярных расширений на краях плоских цистерн. Принято различать в зоне диктиосомы проксимальный или формирующийся, цис-участок, и дистальный или зрелый, транс-участок (рис. 15.5). Между ними располагается средний или промежуточный участок АГ. Во время деления клеток сетчатые формы АГ распадаются до диктиосом, которые пассивно
и случайно распределяются по дочерним клеткам. При росте клеток общее количество диктиосом увеличивается.


Рис. 3. Типы аппарата Гольджи (по Ченцову Ю.С., 2010)
Примечание: а ─ сетчатый в клетках кишечного эпителия; б ─ диффузный в клетках спинального ганглия; I ─ ядро; 2 ─ АГ; 3 ─ ядрышко.
АГ обычно поляризован в секретирующих клетках: его проксимальная часть обращена к цитоплазме и ядру, а дистальная ─ к поверхности клетки. В проксимальном участке к стопкам сближенных цистерн примыкает зона мелких гладких пузырьков и коротких мембранных цистерн.

Рис. 4. Аппарат Гольджи (АГ) в электронном микроскопе(по Ченцову Ю.С., 2010)
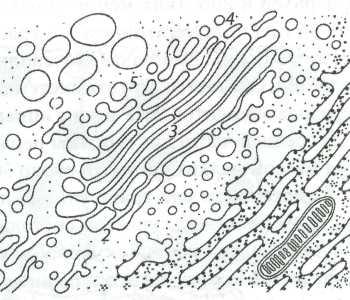
Рис. 5. Схематическое изображение компонентов аппарата Гольджи (по Ченцову Ю.С., 2010)
Примечание: 1 ─ ЭПР-АГ (ERGIC) ─ промежуточная зона; 2 ─ цис-зона, проксимальный участок; 3 ─ медиальный ─ средний участок; 4 ─ транс-дистальный участок; 5 ─ транс-сеть АГ.
В средней части диктиосомы периферия каждой цистерны также сопровождается массой мелких вакуолей около 50 нм в диаметре.
В дистальном или транс-участке диктиосом к последней мембранной плоской цистерне примыкает участок, состоящий из трубчатых элементов и массой мелких вакуолей, часто имеющих фибриллярную опушѐнность по поверхности со стороны цитоплазмы ─ это опушѐнные или окаймленные пузырьки такого же типа, как и окаймленные пузырьки при пиноцитозе (от др.-греч. πίνω ─ пью, впитываю и κύτος ─ вместилище, клетка ─ захват клеточной поверхностью жидкости с содержащимися в ней веществами; процесс поглощения и внутри клеточного разрушения макромолекул).
Это так называемая транс-сеть аппарата Гольджи (TGN), где происходит разделение и сортировка секретируемых продуктов. Еще дистальнее располагается группа более крупных вакуолей ─ это уже продукт слияния мелких вакуолей и образования секреторных вакуолей.
При изучении толстых срезов клеток в мегавольтный электронный микроскоп было найдено, что в клетках отдельные диктосомы могут быть связаны друг с другом системой вакуолей и цистерн. Так что образуется рыхлая трехмерная сеть, выявляемая в световом микроскопе. В случае диффузной формы АГ каждый отдельный его участок представлен диктиосомой. В клетках животных часто с зоной мембран аппарата Гольджи ассоциированы центриоли; между радиально отходящих от них пучков микротрубочек лежат группы стопок мембран и вакуолей, которые концентрически окружают клеточный центр. Эта связь, вероятно, отражает участие микротрубочек в движении вакуолей.
Функции аппарата Гольджи. Наряду с белками в аппарате Гольджи осуществляется транспорт мембранных липидов.
1. Разделение белков на 3 потока:
● Цис-отдел (ближний к ядру); лизосомальный ─ гликозилированные белки (с маннозой) поступают в цис-отдел комплекса Гольджи, некоторые из них фосфорилируются, образуется маркёр лизосомальных ферментов ─ манноза-6-фосфат. В дальнейшем эти фосфорилированные белки не будут подвергаться модификации, а попадут в лизосомы.
● Медиальный отдел; конститутивный экзоцитоз (конститутивная секреция). В этот поток включаются белки и липиды, которые становятся компонентами поверхностного аппарата клетки, в том числе гликокаликса, или же они могут входить в состав внеклеточного матрикса.
● Транс-отдел (самый отдаленный от ядра); индуцируемая секреция ─ сюда попадают белки, которые функционируют за пределами клетки, поверхностного аппарата клетки, во внутренней среде организма. Характерен для секреторных клеток.
2. Формирование слизистых секретов (секреторная функция аппарата Гольджи) ─ гликозамингликанов (мукополисахаридов).
●Мембранные элементы АГ участвуют в сегрегации и накоплении продуктов, синтезированных в ЭПР, участвуют в их химических перестройках, созревании (перестройка олигосахаридных компонентов гликопротеинов в составе водорастворимых секретов или в составе мембран), (рис. 6).
●В цистернах АГ происходит синтез полисахаридов, их взаимосвязь с белками, приводящая к образованию мукопротеидов.
●Главное, с помощью элементов аппарата Гольджи происходит процесс выведения готовых секретов за пределы клетки. Кроме того, АГ ─ источник клеточных лизосом.
●Участие АГ в процессах выведения секреторных продуктов было очень хорошо изучено на примере экзокринных клеток поджелудочной железы. Для этих клеток характерно наличие большого числа секреторных гранул (зимогеновых гранул), которые представляют собой мембранные пузырьки, заполненные белковым содержимым. В составе белков зимогеновых гранул входят разнообразные ферменты: протеазы, липазы, карбогидразы, нуклеазы.
При секреции содержимое этих зимогеновых гранул выбрасывается из клеток в просвет железы, а затем перетекает в полость кишечника. Так как основным продуктом, выводимым клетками поджелудочной железы, является белок, то исследовали последовательность включения радиоактивных амино-кислот в различные участки клетки (рис. 7).
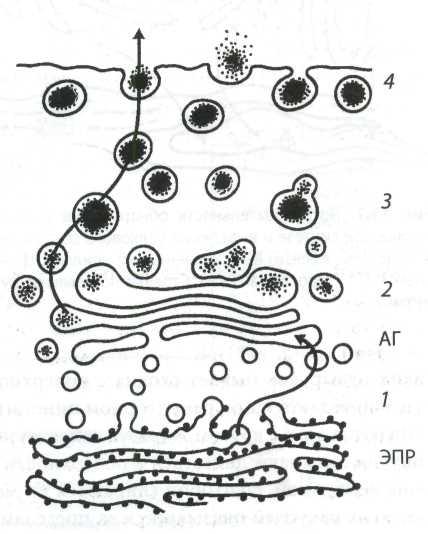
Рис. 6. Схема связи ЭПР, аппарата Гольджи с образованием и выделением зимогена из ацинарных клеток поджелудочной железы (по Ченцову Ю.С., 2010)
Примечание: 1 ─ переходная зона между ЭПР и АГ; 2 ─ зона созревания секреторных гранул; 3 ─ отделившиеся от АГ зимогеновые гранулы; 4 ─ их выход (экзоцитоз) за пределы клетки.
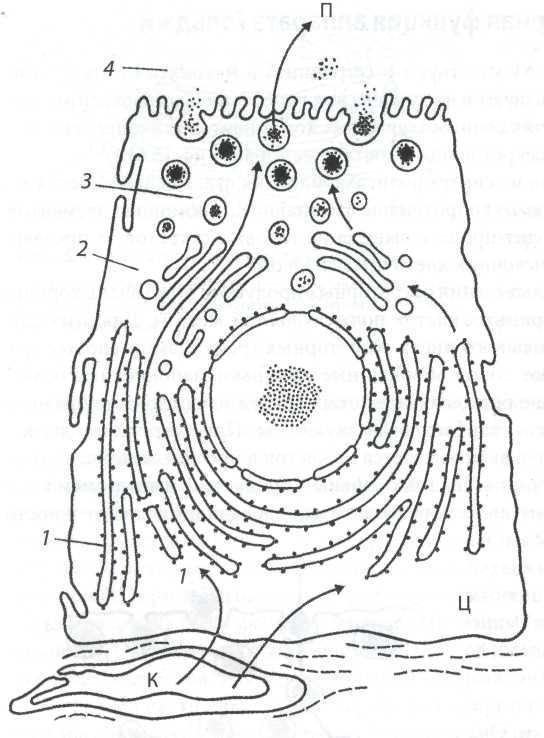
Рис. 7. Последовательность обнаружения [1-4) метки от 3Н-лизина при синтезе и выведении белкового секрета из клетки поджелудочной железы (по Ченцову Ю.С., 2010)
Примечание: К ─ кровеносный апилляр, Ц ─ цитоплазма клетки; П ─ просвет железы. Стрелки показывают пути миграции метки.
Для этого животным вводили меченную тритием аминокислоту (3Н-лейцин) и с помощью электронно-микроскопической радиоавтографии следили во времени за локализацией метки. Оказалось, что АГ является промежуточным звеном между собственно синтезом секретируемого белка и выведением его из клетки. Также подробно процессы синтеза и выведения белков были изучены на других клетках (молочная железа, бокаловидные клетки кишечника, щитовидная железа и др.), и были исследованы морфологические особенности этого процесса. Синтезированный на рибосомах, экспортируемый белок отделяется и накапливается внутри цистерн ЭПР, по которым он транспортируется к зоне мембран АГ. Здесь от гладких участков ЭПР отщепляются мелкие вакуоли, содержащие синтезированный белок, которые поступают в зону вакуолей в проксимальной части диктиосомы (ЭПР-АГ комплекс). В этом месте вакуоли могут сливаться друг с другом и с плоскими цис-цистернами диктиосомы. Таким образом происходит перенесение белкового продукта уже внутри полостей цистерн АГ.
●В аппарате Гольджи происходит не просто «перекачка» продуктов из одной полости в другую, но и постепенно идет их «созревание», модификация белков, которая заканчивается «сортировкой» продуктов, направляющихся или к лизосомам, или к плазматической мембране, или к секреторным вакуолям.
3. Формирование углеводных компонентов гликокаликса ─ в основном, гликолипидов.
4. Сульфатирование углеводных и белковых компонентов гликопротеидов и гликолипидов. В аппарате Гольджи происходит также сульфатирование глюкозаминогликанов и некоторых белков.
5. Частичный протеолиз белков ─ иногда за счет этого неактивный белок переходит в активный (проинсулин превращается в инсулин).
6. Транспорт веществ из эндоплазматической сети. Аппарат Гольджи асимметричен ─ цистерны, располагающиеся ближе к ядру клетки (цис-Гольджи) содержат наименее зрелые белки, к этим цистернам непрерывно присоединяются мембранные пузырьки ─ везикулы, отпочковывающиеся от гранулярного эндоплазматического ретикулума (ЭПР), на мембранах которого и происходит синтез белков рибосомами. Перемещение белков из эндоплазматической сети (ЭПС) в аппарат Гольджи происходит неизбирательно, однако не полностью или неправильно свернутые белки остаются при этом в ЭПС. Возвращение белков из аппарата Гольджи в ЭПС требует наличия специфической сигнальной последовательности (лизин-аспарагин-глутамин-лейцин) и происходит благодаря связыванию этих белков с мембранными рецепторами в цис-Гольджи.
7. Модификация белков в аппарате Гольджи. В цистернах аппарата Гольджи созревают белки предназначенные для секреции, трансмембранные белки плазматической мембраны, белки лизосом и т. д. Созревающие белки последовательно перемещаются по цистернам в органеллы, в которых происходят их модификации ─ гликозилирование и фосфорилирование. При О-гликозилировании к белкам присоединяются сложные сахара через атом кислорода. При фосфорилировании происходит присоединение к белкам остатка ортофосфорной кислоты.
В цис-зону аппарата Гольджи синтезированные в ЭПР белки попадают после первичного гликозилирования и редукции там же нескольких сахаридных остатков. В конечном итоге все белки там имеют одинаковые олигосахаридные цепи, состоящие из двух молекул N-ацетил-глюкозамина, шести молекул маннозы (рис. 8). В цис-цистернах начинается вторичная модификация олигосахаридных цепей и их сортировка на два класса. В результате олигосахариды на гидролитических ферментах, предназначенных для лизосом (олигосахариды, богатые маннозой), фосфорилируются, а олигосахариды других белков, направляемых в секреторные гранулы, или к плазматической мембране, подвергаются сложным превращениям, теряя ряд сахаров и присоединяя галактозу, N-ацетилглюкозамин и сиаловые кислоты. При этом возникает специальный комплекс олигосахаридов. Такие превращения олигосахаридов осуществляются с помощью ферментов ─ гликозилтрансфераз, входящих в состав мембран цистерн аппарата Гольджи.
По мере модификации белков в цистернах аппарата Гольджи, они с помощью мелких вакуолей переносятся от цистерн к цистерне в дистальную часть диктиосомы, пока не достигают трубчатой мембранной сети в трансучастке диктиосомы. В этом участке происходит отщепление мелких пузырьков, содержащих уже зрелый продукт. Цитоплазматическая поверхность таких пузырьков бывает сходна с поверхностью окаймленных пузырьков, которые наблюдаются при рецепторном пиноцитозе. Отделившиеся мелкие пузырьки сливаются друг с другом, образуя секреторные вакуоли. После этого секреторные вакуоли начинают двигаться к поверхности клетки, соприкасаются с плазматической мембраной, с которой сливаются их мембраны, и, таким образом, содержимое этих вакуолей оказывается за пределами клетки. Морфологически этот процесс экструзии (выбрасывания) напоминает пиноцитоз, только с обратной последовательностью стадий. Он носит название экзоцитоз. Экзоцитоз (от греч. Έξω ─ внешний и κύτος ─ клетка) ─ у эукариот клеточный процесс, при котором внутриклеточные везикулы (мембранные пузырьки) сливаются с внешней клеточной мембраной. При экзоцитозе содержимое секреторных везикул (экзоцитозных пузырьков) выделяется наружу, а их мембрана сливается с клеточной мембраной. Практически все макромолекулярные соединения (белки, пептидные гормоны и др.) выделяются из клетки этим способом.
Такое описание событий является только общей схемой участия аппарата Гольджи в секреторных процессах. Дело усложняется тем, что одна и та же клетка может участвовать в синтезе многих выделяемых белков, может их друг от друга изолировать и направлять к клеточной поверхности или же в состав лизосом.
Каждая зона в диктиосомах имеет свой набор ферментов гликозилирования, то гликопротеиды как бы по эстафете переносятся из одного мембранного отсека («этажа» в стопке цистерн диктиосомы) в другой и в каждом подвергаются специфическому воздействию ферментов.

Рис. 8. Пути вторичного гликозилирования белков в аппарате Гольджи (по Ченцову Ю.С., 2010)
Примечание: а ─ первичное гликозилирование в ЭПР; б ─ результат вторичного гликозилирования в АГ белков плазматической мембраны и секреторных вакуолей; в ─ результат вторичного гликозилирования белков лизосом; Ман ─ манноза; Асп ─ аспарагин; Гл ─ глюкоза; СК ─ сиаловая кислота; Га ─ N-ацетилглюкозамин; Гал ─ галактоза.
В цис-участке происходит фосфорилирование манноз в лизосомных ферментах и образуется особая маннозо-6-группировка, характерная для всех гидролитических ферментов, которые потом попадут в лизосомы.
В средней части диктиосом протекает вторичное гликозилирование секреторных белков: дополнительное удаление маннозы и присоединение N-ацетилглюкозамина.
В транс-участке к олигосахаридной цепи присоединяются галактоза и сиаловые кислоты (рис. 9).

Рис. 9. Локализация ферментов модификации белков в аппарате Гольджи (по Ченцову Ю.С., 2010)
Примечание: 1 ─ синтез белка в ЭПР; 2 ─ фосфорилирование лизосомных олигосахаридов; 3 ─ отщепление маннозы; ─ 4присоединение N-ацетилглюкозамина; 5 ─ присоединение маннозы; 6 ─ присоединение сиаловой кислоты; 7 ─ сортировка белков на рецепторах в транссети; 8 ─ лизосома; 9 ─ секреторная вакуоль; 10 ─ плазмалемма
8. Взаимосвязь аппарата Гольджи и эндоплазматического ретикулума. В разных областях аппарата Гольджи имеет место посттрансляционная модификация белков (рис. 9). Созревшие (модифицированные) белки переносятся везикулами в различные отделы клетки, такие, как лизосомы (4), цитоплазматическая мембрана (6) или секреторные пузырьки (5). Последние высвобождают свое содержимое к межклеточное пространство, сливаясь с плазматической мембраной (экзоцитоз). Эти транспортные процессы могут быть конститутивными, т.е. проходить постоянно, или регуляторными, т.е. управляться химическими сигналами. Направленность процесса в первую очередь зависит от сигнальной последовательности синтезируемого белка.
В образцах препаративно выделенных зон АГ при негативном контрастировании видно, что к проксимальной части диктиосомы примыкает сетевидная или губкообразная система мембранных полостей. Считается, что эта система (ЭПР ─ АГ) может представлять собой зону перехода элементов ЭПР в зону аппарата Гольджи.

Рис. 9. Аппарат Гольджи и шероховатый эндоплазматический ретикулум
Эндоплазматический ретикулум (ЭПР) ─ протяженная замкнутая мембранная структура, построенная из сообщающихся трубкообразных полостей и мешочков, называемых цистернами. В области ядра ЭПР сообщается с внешней ядерной мембраной. Между шероховатым и гладким ЭПР имеется морфологическое различие: мембраны шероховатого ЭР усеяны множеством рибосом, в то время как гладкий ЭР не имеет связанных рибосом.
Шероховатый эндоплазматический ретикулум (А) ─ место активного биосинтеза белков. Именно здесь синтезируются белки, которые будут функционировать в составе мембран, лизосом или секретироваться из клетки. Остальные белки синтезируются в цитоплазме на рибосомах, не связанных с мембранами ЭПР. Белки, синтезированные на шероховатом ЭПР (1), претерпевают посттрансляционные модификации (созревание белков). Они либо остаются внутри шероховатого ЭР в виде мембранных белков, либо транспортируются с помощью везикул (2) в аппарат Гольджи (3). Транспортные везикулы образуются почкованием мембран, а затем исчезают, сливаясь с ними.
Гладкий эндоплазматический ретикулум ─ эндоплазматическая сеть, не имеющая связанных рибосом (ГЭР), (рис. 10). Он занимает в клетке сравнительно небольшой объем.

Рис. 10. Аппарат Гольджи и гладкий эндоплазматический ретикулум
Выраженный ГЭР имеется в клетках с активным обменом липидов, таких, как гепатоциты и клетки Лейдига. Для ГЭР характерна замкнутая система разветвленных канальцев. ГЭР принимает участие в синтезе липидов. Биосинтез осуществляется ферментами, закрепленными на мембранах ГЭР. Здесь локализован синтез фосфолипидов и отдельные стадии синтеза холестерина. В ГЭР специализированных клеток эндокринной системы протекают различные стадии синтеза стероидных гормонов. В ГЭР локализованы также процессы метаболической трансформации ксенобиотиков. В этих реакциях принимает участие система цитохрома Р450, которую считают основной системой ГЭР. ГЭР выполняет функцию депо ионов Са2+, поддерживающего низкий уровень Са2+ в цитоплазме. Эта функция более всего свойственна саркоплазматическому ретикулуму, специализированной форме ГЭР мышечных клеток (см. с. 326). В мембранах ГЭР локализованы управляемые Са2+-каналы и энергозависимые Са2+-насосы, а высокая концентрация ионов Са2+ в цистернах поддерживается при участии Са2+-связывающих белков.
9. Сортировка белков в аппарате Гольджи. Через аппарат Гольджи проходит по крайней мере три потока синтезированных клеткой нецитозольных белков: поток гидролитических ферментов в компартмент лизосом, поток выделяемых белков, которые накапливаются в секреторных вакуолях, и выделяются из клетки только по получении специальных сигналов, поток постоянно выделяемых секреторных белков. Существует специальный механизм пространственного разделения этих разных белков и их путей следования.
Аппарат Гольджи в нейронах. Спинномозговой узел котенка. Обработка четырехокисью осмия. Спинномозговые узлы млекопитающих находятся вблизи спинного мозга и содержат чувствительные нервные клетки. При малом увеличении видны крупные нервные клетки ─ нейроны или нейроциты (1) ─ округлой формы, располагающиеся преимущественно в периферических отделах спинномозгового узла.

Рис. 1. Аппарат Гольджи в нейронах. Спинномозговой узел котенка. Обработка четырехокисью осмия
Нейроны ─ наиболее крупные шаровидные клетки с отростками. Однако отростки нейронов выявляются только при специальной гистологической обработке и поэтому на данном препарате не видны. Между скоплениями нейронов находятся нервные волокна (2) и соединительная ткань. Вокруг нейронов видны в виде темных точек ядра (3) клеток-сателлитов или мантийных глиоцитов. Нейроциты отличаются друг от друга размерами, что связано как с действительными различиями их величины, так и направлением плоскости, через которую прошел срез. Надо найти нейроны, в которых видны структурные компоненты, и изучить их при большом увеличении (Б). Вокруг ядра (1) на светлом фоне нейроплазмы (2) выделяется черная извилистая сеть (3). При восстановлении четырехокиси осмия металлический осмий откладывается черным осадком на поверхность структурных компонентов аппарата Гольджи, придавая ему вид клубка нитей или сети (отсюда старое название этой структуры ─ внутриклеточный сетчатый аппарат). Аппарат Гольджи состоит из изогнутых и анастомозирующих между собой нитей и перекладин. Иногда эта сеть вплотную прилегает к ядру и охватывает его, образуя как бы корзинку (4). В других нейронах (5) она располагается несколько отступая от ядра. Встречаются нейроциты, в цитоплазме которых аппарат Гольджи состоит из отдельных палочек, чешуек, фрагментов (6), не связанных между собой и разбросанных по всей нейроплазме. Строение и расположение в цитоплазме аппарата Гольджи связаны с различным функциональным состоянием нейронов. В нервных клетках (7), ядро которых не попало в срез, аппарат Гольджи занимает центральную часть клетки. В поверхностно срезанных нейроцитах (8) он срезан тангенциально; в этом случае он имеет вид сети, занимающей своими перекладинами всю нейроплазму. Изучение аппарата Гольджи на светооптическом уровне свидетельствует, что сетевидная форма этого образования характерна для большинства клеток тканей взрослого организма. Электронная микроскопия показала, что аппарат Гольджи представляет собой сложную мембранную систему, в состав которой входят три типа структур: цистерны, микропузырьки и вакуоли. Эти структурные особенности аппарата Гольджи послужили основанием для его названия ─ комплекс Гольджи или пластинчатый комплекс. Участок цитоплазмы, в котором располагается этот органоид, называют «зоной Гольджи». Анализ ультраструктурных и биохимических особенностей пластинчатого комплекса в различных клетках животных и растений показал, что структура его элементов в разных объектах сходна и он функционально связан с процессами, обеспечивающими жизнедеятельность клетки. Такие наблюдения дают основание для причисления комплекса Гольджи к органоидам общего значения. Этот органоид особенно сильно выражен в клетках с интенсивным синтезом веществ (нервные, яйцевые, железистые). Комплекс Гольджи весьма активный органоид, участвующий в синтезе веществ, обеспечивающих рост и замену отмирающих частей клетки, а также потребности всего организма. Последнее достигается путем выделения синтезируемого продукта за пределы клетки.