

Л_3_Ферменты
.pdfФерменты
Биокатализаторы. Ферменты и рибозимы.
Химический и биологический катализ (общее и различия).
Механизм действия.
Кинетика химических реакций.
Константа Михаэлиса.
Строение и свойства ферментов как белковых молекул.
Принципы регуляции активности ферментов.
Изоферменты.
Классификация и номенклатура ферментов.
Биокатализаторы
Катализаторы — химические вещества, которые ускоряют протекание химической реакции, но не входят в состав ее продуктов (Р).
Количество катализатора, в отличие от других реагентов, после реакции не изменяется.
Катализатор:
участвует в реакции, обеспечивая более быстрый путь
реагирует с исходным веществом
подвергает превращениям получившееся промежуточное соединение
отщепляет продукт
снова реагирует с исходным веществом.
Этот каталитический цикл многократно (до миллиона раз) повторяется.
Схема протекания реакции с катализатором
Катализаторы следует отличать от инициаторов. Например, перекиси распадаются на свободные радикалы, которые могут инициировать радикальные цепные реакции.
Инициаторы расходуются в процессе реакции, поэтому их нельзя считать катализаторами. Ингибиторы иногда ошибочно считают отрицательными катализаторами. Но ингибиторы, например,
цепных радикальных реакций, реагируют со свободными радикалами и, в отличие от катализаторов, не сохраняются. Другие ингибиторы (каталитические яды) связываются с катализатором и его дезактивируют, здесь имеет место подавление катализа, а не отрицательный катализ. Отрицательный катализ в принципе невозможен: он обеспечивал бы для реакции более медленный путь, но реакция, естественно, пойдёт по более быстрому, в данном случае, не катализированному, пути.
Биокатализаторы - химические вещества, катализирующие (ускоряющие) химические реакции в живых системах: ферменты и рибозимы (РНК). Ферменты синтезируются на рибосомах, а РНК - в ядре.
Ферменты – это белки, обладающие каталитической активностью. Название происходит от латинского слова fermentatio - брожение (процесс, связанный с выделением газов).
Энзимы (Е) - это другое название ферментов. Оно происходит от
греческого слова (англ. Enzyme) и означает “в дрожжах”. |
|
||||
Термины |
«фермент» |
и |
«энзим» |
давно |
используют |
как синонимы (первый в основном в |
русской |
и немецкой научной |
|||
литературе, второй — в англо- и франкоязычной). |
|
|
|||
Наука о ферментах называется энзимологией, а не ферментологией
(чтобы не смешивать корни слов латинского и греческого языков). Известно более 4000 ферментов.
Рибозим (сокращение от «рибонуклеиновая кислота» и «энзим») -
это молекула РНК, с каталитическим действием (каталитическая РНК).
До открытия рибозимов ферменты считались единственными органическими катализаторами.
Каталитическая активность РНК впервые была обнаружена в 1980-е годы у пре-рРНК Томасом Чеком, изучавшим сплайсинг РНК, и Сидни Альтманом, работавшим с бактериальной рибонуклеазой P.
В 1989 году Чек и Альтман получили Нобелевскую премию по химии за «обнаружение каталитических свойств РНК».
Действие
Активная часть рибосомы — является рибозимом.
Кофактором некоторых рибозимов могут быть двухвалентные йоны металлов, например Mg+2.
РНК может содержать наследственную информацию.
Это позволило Уолтеру Гилберту выдвинуть предположение, что в древности РНК использовалась в качестве:
генетического материала,
катализаторов
структурных компонентов клетки.
Впоследствии эти роли были перераспределены между ДНК и белками. Эта гипотеза сейчас известна как Гипотеза мира РНК.
Если РНК были первыми молекулярными машинами, которые использовались в ранних живых клетках, то рибозимы, существующие сегодня (например, аппарат рибосомы), могут считаться живыми ископаемыми — образцами живых существ, состоящих из нуклеиновых кислот.
Исследования свертывания прионов показывают, что РНК может катализировать свёртывание белка в патологические конфигурации.
ОБЩЕЕ ДЛЯ ХИМИЧЕСКОГО И БИОЛОГИЧЕСКОГО КАТАЛИЗА
Механизм катализа общий для химических и биологических катализаторов: 1) Ускорение химической реакции за счет снижения энергии активации. Энергии активации (ЭА) – это та часть свободной энергии, которую необходимо придать молекуле субстрата (S), чтобы перевести ее в активное состояние. Только в таком состоянии они вступают в реакцию.
Свободная энергия (G) – это та часть потенциальной энергии реагирующих веществ, которая может быть использована для осуществления полезной работы. В графическом изображении ЭА – это высота энергетического барьера, который надо преодолеть для осуществления химической реакции.
Энергия активации без катализатора существенно выше энергии активации с катализатором.
G (Свободная энергия)
|
Активированное состояние (переходное соединение - |
|
точка дефуркации) |
ЭАбк |
Энергия активации без катализатора (ЭАбк >>>ЭАк) |
|
Энергия активации с катализатором (ЭАк) |
ЭАк |
|
GS |
Исходное состояние (энергия субстрата) |
G |
Стандартное изменение G ( G = GS – GP = const) |
GP |
Конечное состояние (энергия продукта) |
 Ход реакции
Ход реакции
Энергетическая схема химической реакции.
2)Катализаторы ускоряют только спонтанно протекающие реакции, т.е. термодинамически разрешенные. Начать реакцию самостоятельно они не могут.
3)Не влияют на стандартное изменения свободной энергии ( G = GS –
GP = const). В ходе реакции оно не изменяется и служит константой для данной реакции.
4)Не влияют на константу равновесия ( G = RTLnK). Поскольку газовая постоянная (R), абсолютная температура (T) и стандартное изменение свободной энергии величины постоянные, то и натуральный логарифм константы термодинамического равновесия (LnK), а также и сама эта константа (K) должны иметь постоянную величину.
5)Это значит, что катализаторы не сдвигают точку
термодинамического равновесия.
6)Они только повышают скорость достижения системой термодинамического равновесия.
7)Остаются неизменными после реакции. Возможно их повторное использование.
8)Действуют в ничтожно малых концентрациях.
9)Принципы кинетики химической реакции общие, т.е. подчиняются закону действующих масс.

РАЗЛИЧИЯ В ХИМИЧЕСКОМ И БИОЛОГИЧЕСКОМ КАТАЛИЗЕ
КИНЕТИКА ФЕРМЕНТАТИВНОЙ РЕАКЦИИ
Кинетика изучает скорость химической реакции. Любая химическая реакция характеризуется константой термодинамического равновесия:
Кр = k+1/k–1.
Величина обратная Кр называется субстратной константой – КS. Это
константа диссоциации фермент-субстратного комплекса (ES).
Она определяет степень химического сродства фермента к субстрату. Чем ниже значение КS, тем выше это сродство.
Образование фермент-субстратного комплекса отличает биокатализ от химического катализа:
k+1 |
k+2 |
|
Е + S |
ES |
E + P |
k–1 |
|
|
Количественное соотношение между концентрацией субстрата и скоростью ферментативной реакции устанавливает уравнение Бригса-
Холдейна:
|
Vmax · [S] |
|
v = |
|
(1) |
|
||
|
Km + [S] |
|
Если v = Vmax/2, то Km = [S]. Следовательно, константа Михаэлиса
(Km) – это такая концентрация субстрата, при которой скорость
ферментативной реакции (v) достигает половины от максимальной
(Vmax).
Она характеризует степень химического сродства фермента к субстрату так же, как КS.
При этом учитывается влияние образующегося продукта на скорость реакции. В этом состоит отличие Km от КS.
Константу Михаэлиса можно рассчитать экспериментально по уравнению, в числителе которого стоит сумма констант скоростей распада ES на исходные и конечные компоненты ферментативной реакции:
k–1 + k+2 |
k–1 |
k+2 |
k+2 |
||||
Km = |
|
= |
|
+ |
|
= КS + |
|
|
|
|
|
||||
k+1 |
k+1 |
k+1 |
k+1 |
||||
Поскольку Кs = k-1/k+1 , постольку константа Михаэлиса всегда больше константы диссоциации фермент-субстратного комплекса на величину, равную k+2/k+1.
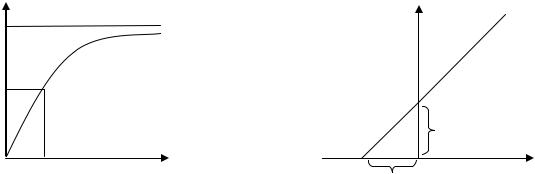
Графический способ расчета Km по методу двойных обратных величин наиболее удобный.
Уравнение Брикса-Холдейна (1) преобразуют в уравнение Лайнуивера-Бэрка (2) путем получения обратных величин. Если обе части уравнения делят или умножают на одно и то же число, то равенство не нарушается:
1/v = Km/ Vmax∙ 1/[S] + 1/Vmax (2)
В итоге гиперболическая зависимость между прямыми величинами начальных скоростей (v) ферментативной реакции и концентрации субстрата - [S], заменяется прямой линией (y = ax + b), как график в координатах
обратных величин (1/v и 1/[S]).
Отсекаемые этой прямой отрезки на оси абсцисс (-1/Km) и ординат (1/Vmax) позволяют рассчитать значения прямых величин Km и Vmax, соответственно.
График уравнения Бригса-Холдейна – это кривая насыщения химической реакции, иллюстрирующая соотношение между концентрацией
субстрата [S] и скоростью реакции v. |
|
|
|
|
|
v |
|
|
1/v |
|
|
Vmax |
|
|
|
|
|
|
|
Km |
|
||
Vmax |
|
Наклон = |
|
|
|
|
|
|
|||
2 |
|
Vmax |
|
||
|
|
|
|
|
1/Vmax |
Km |
[S] |
|
-1/Km |
1/[S] |
|
График уравнения Бригса-Холдейна |
График уравнения Лайнуивера-Бэрка |
||||
Отклонения от этих правил свойственны для аллостерических
ферментов и изоферментов.
Km определяют для характеристики изоферментов, а также для выяснения механизмов действия эффекторов на активность ферментов.
ОСОБЕННОСТИ СТРОЕНИЯ ФЕРМЕНТОВ
Не все белки проявляют каталитические свойства, но все ферменты
– это белки. Белковая природа определяет основные различия в строении и свойствах биологических и неорганических катализаторов:
1. Выделяют две группы ферментов: простые и сложные. Простетическую (небелковую) часть сложных ферментов называют коферментом. Простетическая группа – это главное отличие сложных ферментов от простых.
2. Характерно наличие первичной, вторичной, третичной и четвертичной структуры. Некоторые Е формируют мультиферментные
комплексы (пируватдегидрогеназа) и ансамбли (синтаза ВЖК).
Могут быть структурно связанными с биомембранами
(аденилатциклаза, тирозинкиназа) и c клеточными органеллами:
лизосомы, митохондрии (цепь дыхательных ферментов).
Олигомерные ферменты состоят из нескольких полипептидных цепей (протомеров). Сочетание двух типов протомеров у ЛДГ (тетрамере) дает 5 изоферментов.
3. Все ферменты имеют активный центр. Это главное условие специфичности действия фермента. Он образуется на уровне третичной или четвертичной структуры в виде уникального сочетания радикалов АМК в пространстве.
Различают связывающий участок, распознающий и удерживающий субстрат, и каталитический. Он преобразует субстрат.
Кактивному центру может присоединяться простетическая группа (кофермент). В результате этого из апофермента (белка) и кофермента образуется холофермент - сложный белок, обладающий каталитической активностью.
В качестве коферментов могут быть:
 йоны металлов (1/3 всех коферментов)
йоны металлов (1/3 всех коферментов)
 производные витаминов
производные витаминов
 производные нуклеотидов (АТФ, ЦТФ, УТФ)
производные нуклеотидов (АТФ, ЦТФ, УТФ)
 липоевая кислота
липоевая кислота
 т-РНК
т-РНК
 глутатион и др. вещества.
глутатион и др. вещества.
Каждый фермент имеет один или несколько активных центров, с которыми связывается субстрат.
Эти центры высокоспецифичны, т.е. «узнают» только «свой» субстрат или близкородственные соединения.
Особенность ферментативного катализа состоит в том, что фермент специфическим образом связывает субстрат, и все реакции протекают внутри фермент-субстратного комплекса.
Активный центр формируется из фрагментов полипептидной цепи, в том числе из отдельных аминокислот, содержащих разнообразные функциональные группы. Некоторые из них принимают участие в сорбции субстрата на ферменте, другие - катализируют его химическое превращение.
Ккаталитически активным радикалам белка относятся:
oнуклеофильные группы (имидазол гистидина, оксигруппа серина или
тирозина, тиольная группа цистеина, ε-аминогруппа лизина,
ионизированные карбоксилы аспарагиновой и глутаминовой кислот и др.)
o электрофилы (ион имидазолия, нейонизованные карбоксильные группы, ионы металлов и т. п.).
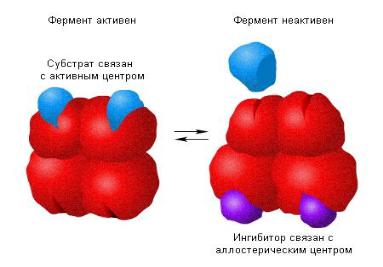
4. Некоторые ферменты имеют аллостерический (регуляторный) центр. Он расположен отдельно от активного центра и связывает аллостерические эффекторы.
Они изменяют пространственную конфигурацию ферментов, в том числе и активный центр, вызывая повышение или понижение
каталитической активности.
В переводе с греческого языка allos означает “другой”, а steros –
пространственный, структурный.
Модель изменения молекулы фермента под действием аллостерического ингибитора
5.Домены ферментов способны:
Присоединять и удерживать в активном центре пиридиновых
дегидрогеназ коферменты: НАД и НАДФ.
Встраиваться в биомембрану в качестве гидрофобного «якоря»:
интегральные, мембранные ферменты.
Выполнять рецепторные функции, располагаясь с внешней стороны мембраны: гидрофильные домены тирозинкиназы.
Катализировать фосфорилирование белков – каталитическая (гидрофильная) часть тирозинкиназы.
ОСНОВНЫЕ СВОЙСТВА ФЕРМЕНТОВ
1. Термолабильность (чувствительность к температуре)
проявляется при нагревании выше 45-50 0С.
До этого значения биокатализ подчиняется теории химической кинетики и закону Вант-Гоффа, когда действует температурный коэффициент (Q10): повышение температуры на 10 0С увеличивает скорость химической реакции в 2-3 раза.
Ферменты функционируют при умеренной температуре и нормальном давлении. В противном случае денатурация приводит к потере
каталитических свойств. |
|
|
|
v |
|
|
|
|
|
v1 |
= f (t 0С) |
|
|
температурный оптимум |
|
|
|
v2 |
= f (денатурации, %) |
0 |
50 |
100 |
t, 0С |
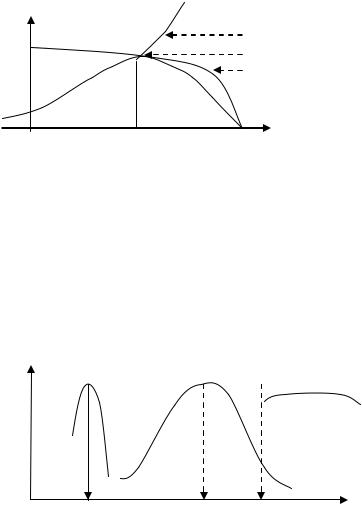
Зависимость скорости ферментативной реакции от температуры.
Форма графика – это результирующая двух зависимостей v1 и v2 .
2. Амфотерность ферментов как полиэлектролитов определяют
йонногенные группы в радикалах АМК: карбоксильная (глу, асп), аминогруппа (лиз), имидазольная (гис) и гуанидиновая (арг).
Они могут участвовать в формировании активного центра Е. Такие же группы могут быть в составе субстратов и коферментов. Это делает скорость ферментативной реакции зависимой от рН:
v пепсин (1) сахараза (2) трипсин (3)
0 |
2 |
4 |
6 |
8 |
10 |
рН |
|
рН1 |
|
рН2 |
рН3 |
|
|
Зависимость скорости ферментативной реакции от рН среды.
Для каждого фермента существуют оптимальные значения рН среды
(рН1, рН2, рН3).
Пики графиков могут быть острыми (1), колоколообразными (2), широкими (3).
Плавность переходов восходящих в нисходящие части графиков, по аналогии с кривыми титрования, получается в результате постепенного изменения баланса положительно и отрицательно заряженных йонногенных групп в активном центре Е. Это ведет к постепенному изменению числа
активных молекул Е в диапазоне рН оптимума.
Свойства полиэлектролитов позволяют использовать методы
электрофореза, йоннообменной хроматографии и изотахофореза
(осаждение белка в изоэлектрической точке) для разделения ферментов на
индивидуальные фракции.
3. Как высокомолекулярные полимеры ферменты образуют
коллоидные растворы и могут подвергаться ультрацентрифугированию, диализу, гельфильтрации и высаливанию.
4. Изоферменты – это множественные формы белков, катализирующих одну и ту же реакцию.
Они различаются по:
физико-химическим свойствам (электрофоретической подвижности)
сродству к субстрату (Кm - различны).
Улактатдегидрогеназы и креатинфосфокиназы определение изоформ путем электрофореза используют в клинической диагностике инфаркта миокарда.
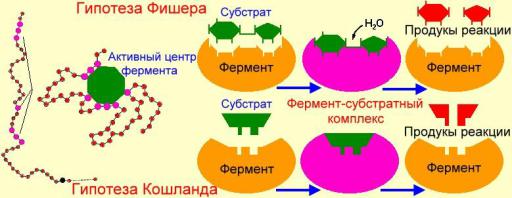
5. Явление индуцированного соответствия Е и S (Кошланд)
указывает на необходимость образования ЕS комплекса.
Это приводит к конформационным изменениям в структуре Е и S.
Субстрат превращается в продукт и комплекс распадается с образованием
свободного Е и продукта реакции.
Ранее предполагали, что активный центр фермента связывается с субстратом по принципу «ключ-замок»: гипотеза Фишера.
Появление в системе молекулы фермента и субстрата вызывает:
притягивание друг к другу под действием электростатических сил
ориентацию молекулы субстрата в активном центре фермента
молекулярное узнавание.
Взависимости от степени совпадения формы и соответствия полярности активного центра и субстрата возникает большее или меньшее количество:
водородных связей
точек ван-дер-ваальсового притяжения
точечных ионных и гидрофобных взаимодействий.
Это удерживает молекулу субстрата в активном центре фермента. Образуется фермент-субстратный комплекс. Потом фермент под действием молекулы субстрата и внешних условий меняет свою конформацию и, как следствие, форму активного центра.
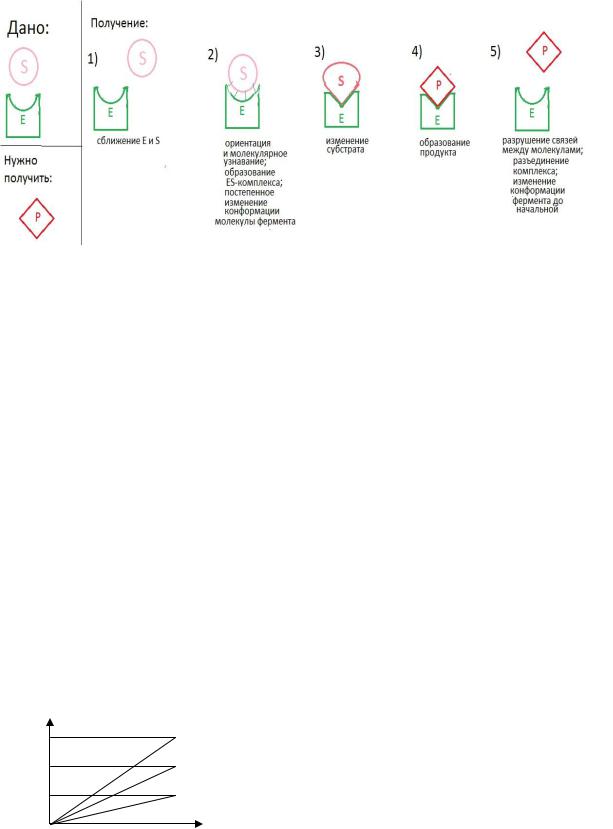
При этом создаются условия для изменения субстрата. В конце концов, образуется продукт и водородные связи разрушаются. Продукт удаляется из активного центра, а сам фермент постепенно возвращает свою изначальную конформацию, и вновь готов превращать молекулы субстрата.
Схема процесса индуцированного соответствия Е и S (Кошланд)
6. По эффективности действия Е значительно превосходят неорганические катализаторы. Моль йонов Fe+2 при O 0C в течение 1 c разлагает 10-5 молей Н2О2. Соответствующее количество каталазы в тех же условиях – 10+5 молей.
7.Субстратная специфичность бывает нескольких типов:
относительная (групповая) - ферменты действуют на определенный тип связей (протеиназы, эстеразы)
абсолютную специфичность проявляют ферменты, расщепляющие
единственный субстрат: аргиназа – аргинин, сахараза - сахарозу
некоторые ферменты действуют на уровне оптических изомеров
(стереоспецифичность): оксидазы L- и D-АМК, α- и β-гликозидазы.
8.Способность к регуляции – это наиболее существенное отличие биологических от неорганических катализаторов. Скорость ферментативной реакции зависит от концентрации субстрата (насыщение и ингибирование избытком), от температуры (инактивация при высоких и сохранение каталитических свойств при низких значениях), от рН среды и концентрации фермента – [Е]. При избытке [S] эта зависимость прямопропорциональная:
Vmax |
|
|
V3 |
[E3] |
Если [E1] ‹ [E2] ‹ [E3] , то V1 ‹V2 ‹V3 |
V2 |
[E2] |
|
V1 |
[E1] |
|
Время Зависимость скорости реакции от концентрации фермента.