

Структура днк
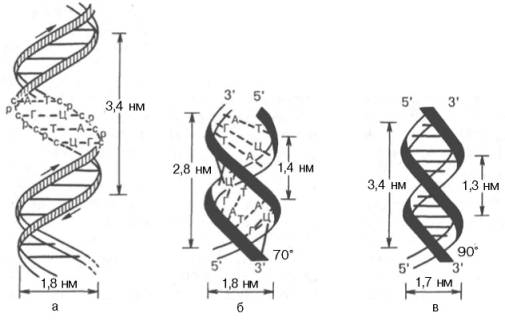
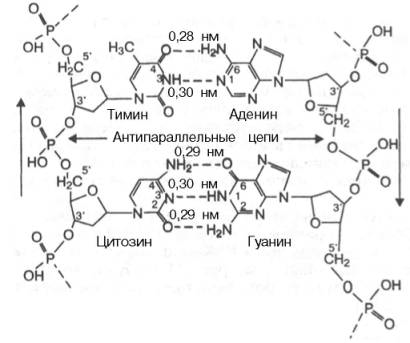
Принцип комплементарности.
Помимо ДНК в клетках встречаются три разновидности РНК: информационные
(и-РНК), транспортные (т-РНК) и рибосомные (р-РНК). Все они отличаются от ДНК рядом особенностей. Во-первых, вместо азотистого основания тимина они содержат урацил. Во-вторых, вместо сахара дезоксирибозы они содержат рибозу. В-третьих, они, как правило, односпиральные.
Переваривание и всасывание нуклеиновых кислот. Катаболизм.
С пищей в сутки в организм поступает около 1 г нуклеиновых кислот.
Переваривание нуклеиновых кислот происходит в тонком кишечнике. Сначала, поступившие с пищей нуклеиновые кислоты под действием ферментов панкреатического сока – нуклеаз – превращаются в мононуклеотиды. Затем уже под влиянием ферментов тонкого кишечника от мононуклеотидов отщепляется фосфорная кислота, и образуются нуклеозиды. Часть нуклеозидов расщепляется затем на азотистое основание и углевод.
Продукты переваривания нуклеиновых кислот поступают в кровь, а затем в печень и другие органы.
В клетках организмов обмен РНК протекает значительно более интенсивно, чем обмен ДНК. В конечном итоге нуклеиновые кислоты расщепляются на азотистые основания, углеводы и фосфорную кислоту.
Далее пуриновые азотистые основания в процессе катаболизма теряют аминогруппу в виде аммиака, окисляются и превращаются в мочевую кислоту.
Пиримидиновые основания подвергаются более глубокому расщеплению до углекислого газа, воды и аммиака.
Углеводы вовлекаются в ГМФ-путь распада и превращаются в глюкозу.
Фосфорная кислота распаду не подвергается. Она используется в реакциях фосфорилирования и фосфолиза или при избытке выделяется из организма с мочой.
Синтез нуклеотидов.
Все клетки организма способны синтезировать необходимые нуклеиновые кислоты и не нуждаются в наличии в пище готовых нуклеиновых кислот или их составных частей. Поэтому содержание готовых нуклеиновых кислот в пище для организма принципиального значения не имеет, хотя продукты их распада могут частично использоваться организмом.
Синтез пуриновых и пиримидиновых нуклеотидов происходит на основе рибозо-5-фосфата. из глюкозы при её распаде по ГМФ-пути. Свободные азотистые основания обычно для этого синтеза не используются.
При синтезе пуриновых нуклеотидов к рибозо-5-фосфату присоединяются атомы углерода и азота, из которых образуется пуриновое кольцо. Источниками этих атомов являются аминокислоты глицин, глутамин, аспарагиновая кислота. Часть атомов углерода поставляется коферментами, содержащими в своём составе фолиевую кислоту и биотин. Промежуточным продуктом синтеза пуриновых нуклеотидов является инозиновая кислота. Далее из инозиновой кислоты образуются пуриновые нуклеотиды.
Синтезу пиримидиновых нуклеотидов предшествует образование необычного азотистого основания оротовой кислоты, содержащей пиримидиновое кольцо. Синтезируется оротовая кислота из аммиака и аспарагиновой кислоты. Оротовая кислота присоединяется к рибозо-5-фосфату и возникает пиримидиновый нуклеотид оротидинмонофосфат. Далее оротовая кислота в составе этого нуклеотида преобразуется в обычные азотистые основания, в результате чего появляются пиримидиновые нуклеотиды.
В связи с высокой важностью оротовой кислоты в спортивной практике в качестве пищевой добавки используется её соль оротат калия.
Дезоксирибонуклеотиды образуются из соответствующих рибонуклеотидов путём восстановления входящей в них рибозы в дезоксирибозу.
Синтез нуклеиновых кислот.
Для синтеза нуклеиновых кислот используются мононуклеотиды обязательно в трифосфатной форме. Такие нуклеотиды содержат в своей молекуле три остатка фосфорной кислоты и обладают повышенным запасом энергии. Переход нуклеотидов в трифосфатную форму осуществляется путем взаимодействия с АТФ. Для синтеза РНК используются АТФ, ГТФ, УТФ, ЦТФ. А для синтеза ДНК, соответственно, дАТФ, дГТФ, дТТФ, дЦТФ.
Процесс репликации или редупликации ДНК иначе можно назвать удвоением. Он происходит перед делением клетки. Осуществляет его специальный фермент ДНК полимераза. Этот фермент разделяет две цепи двойной спирали и достраивает к каждой из них комплементарную ей цепь. Таким образом, из одной молекулы образуется две одинаковые дочерние молекулы, причем обе цепи ДНК служат матрицами для дочерних цепей. По мере присоединения к матрице нуклеотиды связываются в полинуклеотидные нити, которые сразу же закручиваются в двойную спираль. Биологический смысл репликации состоит в том, что из одной молекулы ДНК появляются две ее полные копии. Процесс этот идет с очень высокой точностью – ошибки крайне редки.
Процесс синтеза РНК называется транскрипцией. Процесс образования белков на матрицах информационной РНК называется трансляцией.
Транскрипцию осуществляет фермент РНК-полимераза. Этот фермент соединяет между собой рибонуклеотиды, составляющие остов молекулы РНК. Делает это фермент на основании считывания последовательности молекулы ДНК и, достраивая комплементарные ей последовательности. Показано, что в этом процессе только одна из двух цепей ДНК играет роль матрицы. Бывают, правда, и исключения – это ДНК некоторых вирусов. В процессе транскрипции участвует только ограниченный участок ДНК. Именно этот участок ДНК и понимают в молекулярной биологии, как ген.