

5.4. Пути обезвреживания аммиака
Аммиак образуется из аминокислот при распаде других азотсодержащих соединений (биогенных аминов, НУКЛЕОТИДОВ). Значительная часть аммиака образуется в толстой кишке при гниении. Он всасывается в кровь системы воротной вены, здесь концентрация аммиака больше, чем в общем кровотоке.
Аммиак образуется в различных тканях. Концентрация его в крови незначительна, т.к. он является токсичным веществом (0,4 - 0,7мг/л). Особенно выраженное токсическое действие он оказывает на нервные клетки, поэтому значительное его повышение приводит к серьёзным нарушениям обменных процессов в нервной ткани.
ПУТИ ОБЕЗВРЕЖИВАНИЯ АММИАКА.
1. образование АМИДОВ

![]()
Рис. Образование глутамина
ГЛУТАМИН и АСПАРАГИН - нетоксические вещества. Их называют транспортной формой аммиака в организме. Они не проникают через мембраны и в почках распадаются до аминокислот и аммиака.
2. Восстановительное АМИНИРОВАНИЕ альфа – кетоглутаровой кислоты
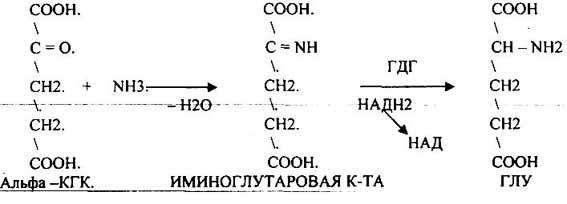
![]()
3. Образование солей АММОНИЯ
![]()
![]()
4. Синтез мочевины - основной путь обезвреживания аммиака - ОРНИТИНОВЫЙ ЦИКЛ.

![]()
АРГИНАЗА обладает абсолютной специфичностью и содержится только в печени. В составе мочевины содержится два атома азота: один поступает из аммиака, а другой выводится из АСП.
Образование мочевины идёт только в печени.
Две первые реакции цикла (образование ЦИТРУЛЛИНА и АРГИНИНОСУКЦИНАТА) идут в МИТОХОНДРИЯХ, остальные в цитоплазме.
В организме в сутки образуется 25г мочевины. Этот показатель характеризует мочевинообразовательную функцию печени. Мочевина из печени поступает в почки, где и выводится из организма, как конечный продукт азотистого обмена.
6.1. Сигнальные молекулы
6.2. Гормоны гипоталамуса
6.3. ГОРМОНЫ ГИПОФИЗА
6.4. ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
6.5. ГОРМОНЫ ПАРАЩИТОВИДНЫХ ЖЕЛЕЗ
6.6. Гормоны половых желез
6.7. Гормоны надпочечников
6.8. Гормоны поджелудочной железы
|
Предыдущий раздел |
Раздел верхнего уровня |
Следующий раздел |
![]()
6. Регуляция обмена веществ
СИГНАЛЬНЫЕ МОЛЕКУЛЫ.
Основные задачи регуляции метаболизма и клеточных функций:
1. внутриклеточное и межклеточное согласование обменных процессов;
2. исключение «холостых» циклов метаболизма, продукты которых не востребованы;
3. эффективное образование и использование энергии;
4. поддержание гомеостаза;
5. приспособление организма к условиям окружающей среды.
Выделяют 2 вида регуляции метаболизма: внутренняя и внешняя.
В случае внутренней регуляции управляющие сигналы образуются и действуют внутри одной и той же клетки (саморегуляция). Внутреня регуляция обеспечивается аллостерическими ферментами, активность которых изменяется при изменении концентрации метаболитов в клетке.
В случае внешней регуляции - управляющие сигналы поступают к клетке из внешней среды.
Внешняя регуляция обеспечивается сигнальными молекулами. Сигнальные молекулы - эндогенные химические соединения, которые в результате взаимодействия с рецепторами, обеспечивают внешнее управление биохимическими процессами в клетках-мишенях.
Клетка-мишень - это клетка, имеющая рецепторы для данного вида сигнальных молекул. Сигнальные молекулы являются лигандами для рецепторов клеток-мишеней.
Характерные особенности сигнальных молекул:
1. малый период жизни (обеспечивает динамичность, оперативность регуляции);
2. высокая биологическая активность (действие развивается при очень низких концентрациях);
3. уникальность, неповторимость действия. Эффекты одного типа сигнальных молекул не могут быть смоделированы другим. Это обеспечивает разнообразие регуляции;
4. наличие эффекта усиления (одна молекула инсулина активирует десятки белков-транспортеров глюкозы);
5. один вид сигнальных молекул может иметь несколько клеток-мишеней (для адреналина рецепторы находятся на мембранах мыщечных и жировых клеток);
6. реакция разных клеток-мишеней на одну и ту же сигнальную молекулу отличается.
Способы внешнего управления клетками-мишенями:
1. Управление экспрессией генов (управление количеством ферментов);
2. Управление активностью ранее синтезированных ферментов;
3. Сочетание этих двух способов.
Виды регуляторных эффектов сигнальных молекул:
1.Эндокринный. Сигнальные молекулы, синтезируемые в железах внутренней секреции, поступают с током крови к клеткам-мишеням. Так действует большинство гормонов.
2.Паракринный - сигнальные молекулы вырабатывают в пределах одного органа или участка ткани. Таким образом действуют большинство факторов роста.
3.Аутокринное - сигнальные молекулы действуют на клетку, их образовавшую.
КЛАССИФИКАЦИЯ СИГНАЛЬНЫХ МОЛЕКУЛ.
1)По химической природе:
1.Органические (производные аминокислот, липидов, простые и сложные белки).
2. Неорганические (оксид азота II - NO).
2)По физико-химическим свойствам:
1.Липофобные - не могут проникать через мембрану клетки. Они растворимы в воде.
2.Липофильные - растворяются в жирах. Свободно проникают через мембрану клетки и действуют через внутриклеточные рецепторы клетки. Например, производные холестерола: минерало -, глюкокортикоиды, эстрогены, андрогены.
3)По биологическому принципу:
1.Гормоны - сигнальные молекулы с выраженным эндокринным эффектом.
2.Цитокины и факторы роста. Это сигнальные молекулы, которые выделяются неспециализированными клетками организма. Они регулируют рост, дифференцировку, пролиферацию соседних клеток. Действие пара- и/или аутокринно.
3.Нейромедиаторы - сигнальные молекулы, вырабатывающиеся нервными клетками, влияющие на обмен веществ и функции иннервируемых тканей. Их действие связано с влиянием на ионные каналы. Они изменяют их проницаемость и вызывают деполяризацию мембраны.
МЕХАНИЗМЫ ДЕЙСТВИЯ СИГНАЛЬНЫХ МОЛЕКУЛ.
Механизм действия органических ЛИПОФИЛЬНЫХ сигнальных молекул характеризуется:
1.взаимодействием сигнальной молекулы с внутриклеточным рецептором;
2.регуляторный эффект связан с изменением количества белка в результате влияния на экспрессию генов,
3. биологическое действие продолжительное, но развивается медленно в пределах нескольких часов.
Факторы, необходимые для их действия:
- сигнальная молекула,
- воспринимающий внутриклеточный рецептор, связанный с шапероном.
- участок ДНК, регулирующий транскрипцию определённых генов (энхансер, сайленсер),
- белоксинтетический аппарат клетки.
Этапы действия липофильных сигнальных молекул:
1. проникновение сигнальной молекулы внутрь клетки,
2. связывание с внутриклеточным рецептором,
3. освобождение шаперона,
4. взаимодействие комплекса сигнальных молекул с регуляторными элементами ДНК, изменение биосинтеза некоторых белков, в том числе и их ферментов.
5. изменение метаболизма и клеточных функций;
Прекращение эффекта за счет инактивации комплекса сигнальная молекула рецептор, протеолиза белков – продуктов регулируемого гена, изменения конформации белков и рецепторов, разрушения мРНК.
Механизм действия ЛИПОФОБНЫХ сигнальных молекул:
1. взаимодействие с поверхностным рецептором,
2. сигнал передаётся от рецептора внутрь клетки (трансдукция) и устанавливается с помощью внутриклеточных регуляторов: высоко- и низкомолекулярных.
Высокомолекулярные регуляторы - это регуляторные белки. Они опосредуют действие сигнальной молекулы внутри клетки.
Низкомолекулярные регуляторы небелковой природы. Его называют второй МЕССЕНДЖЕР (первый МЕССЕНДЖЕР - сама сигнальная молекула. Это ионы кальция, диацилглицерол, инозитол-3-фосфат, цАМФ и цГМФ.
3. биологическое действие обусловлено сочетанием регуляции активности ранее синтезированных белков и регуляция экспрессии генов. Регуляторный эффект двухфазный:
- первая фаза быстрая, но не продолжительная, она обеспечивает изменение структуры и активности ферментов;
- вторая фаза медленная за счёт изменения количества ферментов.
МЕХАНИЗМ ДЕЙСТВИЯ, ЗАВИСИМЫЙ ОТ цАМФ.
Факторы, необходимые для этого:
1. растворимая в воде сигнальная молекула;
2. поверхностные рецепторы клетки-мишени;
3. внутриклеточный трансдуктор G-белок. Состоит из 3 единиц: альфа, бета, гамма. При взаимодействии сигнальной молекулы с рецептором он активируется, его активность обусловлена альфа единицей. Она оказывает влияние на внутриклеточный фермент - АДЕНИЛАТЦИКЛАЗУ (активирует или ингибирует фермент). G-белок способен присоединять ГДФ или ГТФ. Альфа-единица активна, когда связана с ГТФ.
4. АДЕНИЛАТЦИКЛАЗА(АЦ);
5. ПРОТЕИНКИНАЗА-А цАМФ-зависимая. Она катализирует реакцию фосфорилирования белков. В результате белки изменяют активность;
6. Регуляторные элементы ДНК (ЭНХАНСЕР и САЙЛЕНСЕР);
7. ФОСФОДИЭСТЕРАЗА - разрушает ЦАМФ;
8. ФОСФАТАЗА - дефосфорилирует белки;
9. Белково-синтетический аппарат клетки.
Этапы стимулирующего цАМФ - зависимого механизма:
1. взаимодействие сигнальной молекулы с рецептором;
2. изменение конформации G-белка;
3. замена ГДФ на ГТФ в альфа-S единице G-белка;
4. альфа-субъединица (стимулирующая) активирует аденилатциклазу;
5. аденилатциклаза синтезирует цАМФ;
6. цАМФ активирует ПРОТЕИНКИНАЗУ А (ПКА);
7. ПРОТЕИНКИНАЗА А фосфорилирует белки и белковые факторы транскрипции, изменяющие активность и количество ферментов;
8. Прекращение действия, если рецептор освободился
- ФОСФОДИЭСТЕРАЗА - разрушает цАМФ.
- ФОСФАТАЗА - ДЕФОСФОРИЛИРУЕТ белки.
Этапы ингибирующего цАМФ - зависимого механизма:
1. взаимодействие сигнальной молекулы с рецептором;
2. изменение конформации G-белка;
3. замена ГДФ на ГТФ в альфа-S единице G-белка;
4. альфа-субъединица (ингибирующая) угнетает аденилатциклазу;
5. прекращение эффектов цАМФ в клетке
МЕХАНИЗМ ДЕЙСТВИЯ, ЗАВИСИМЫЙ ОТ цГМФ.
Необходимые компоненты:
1. Поверхностный рецептор;
2. Гуанилатциклаза (превращает ГТФ в цГМФ;
3. Протеинкиназа G;
4. Фосфодиэстераза;
5. Фосфатаза.
Рецептор встроен в мембрану клетки и связан с ферментом гуанилатциклазой. При присоединении сигнальной молекулы к рецептору гаунилатциклаза активируется и ускоряет образование цГМФ. Последний активирует протеинкиназу G, он запускает реакцию фосфорилирования белков (ферментов и факторов транскрипции).
ТИРОЗИНКИНАЗНЫЙ МЕХАНИЗМ ДЕЙСТВИЯ.
По данному механизму действуют ферменты, обладающие внутренней тирозинкиназной активностью - способность присоединять фосфат по остаткам тирозина. После взаимодействия мономерного компонента тирозинкиназного рецептора с сигнальной молекулой он димеризуется. внутрення часть рецептора самофосфорилируется. Это вызывает активацию внутренних сигнальных путей. Сигнал передается в ядро, что вызывает изменение роста и дифференцировки. В некоторых клетках после связывания с сигнальной молекулой рецептор интернализуется (погружается внутрь) и проникает в ядро и вызывает описанный ранее эффект.
МЕХАНИЗМ ДЕЙСТВИЯ ОКСИДА АЗОТА (II)
ОКСИД АЗОТА (II) проникает через мембрану клетки, взаимодействует с растворимой гуанилатциклазой. Фермент активируется, в результате чего образуется цГМФ, который увеличивает активность протеинкиназы G, что вызывает соответствующий клеточный ответ.
ОКСИД АЗОТА (II) – молекула, которая регулирует тонус сосудов и апоптоз.