

Глава 2
ПРОДУЦИРОВАНИЕ ОРГАНИЧЕСКОГО ВЕЩЕСТВА В ГЕОСИСТЕМАХ
Общие закономерности развития живого вещества на Земле
Жизнь как совокупность живых организмов на Земле развивается непрерывно более 3 млрд. лет. Это возможно благодаря постоянному притоку солнечной энергии и обмену веществ между организмами и средой. Жизнь в сущности есть функция двух переменных - космической энергии Солнца и глубинной энергии планеты (Ронов, 1978). Все жизненные процессы в организмах совершаются с использованием трансформированной солнечной энергии. Жизнь не может быть сведена к превращениям энергии, но обязательно ими сопровождается (Хильми, 1966).
Жизнь на Земле основана на углеродистых соединениях, причем запасы углерода в равновесной динамической системе атмосфера - океан - живое вещество ограничены, и в то же время некоторая его часть постоянно выводится из системы и подвергается захоронению среди осадочных пород. В этих условиях энергия Земли в аспекте геологической истории выполняет чрезвычайно важную роль, так как при ее посредстве из мантии наряду с другими веществами выделяются летучие углеродистые соединения (главным образом диоксид углерода), пополняющие естественную убыль углерода в атмосфере и гидросфере, у
В ледниковые эпохи льды уничтожили почти всю массу живого вещества на территории своего распространения и одновременно в результате общепланетарного ухудшения климатических условий (снижение среднегодовой температуры воздуха Земли) уменьшилась «плотность жизни» и за пределами материкового оледенения вплоть до экватора. Все это приводило в эпохи максимального одновременного распространения льдов к сокращению массы живого вещества Земли примерно на 20% по сравнению с межледниковьями.
В современную эпоху, особенно за последние 300 лет, имело место существенное сокращение живого вещества Земли. Это происходило в результате: а) почти полного уничтожения живого вещества на площади, занятой строениями, дорогами и карьерами,— около 1,3 млн. км2 (Рябчиков, 1972); б) замены части лесных природных комплексов сельскохозяйственными землями (пашней, сенокосами, пастбищами); в) уменьшения живого вещества лесов по причине недостаточно рациональной их эксплуатации. Вследствие антропогенного воздействия количество живого вещества Земли сократилось до 1,8-1012—1,85-1012 т сухой массы (Рябчиков, 1972), т. е. примерно на 1/4 своей величины доагрикультурного периода, и приблизилось к уровню, характерному для ледниковых эпох плейстоцена.
Наряду с «плотностью жизни» не менее важным показателем развития природного комплекса выступает его первичная биологическая продукция — количество органического вещества, создаваемого фотосинтезирующими растениями в течение года в расчете на единицу площади. Последний показатель отражает интенсивность развития жизни, так как энергия, заключенная в продукции растительности, в дренированных комплексах почти полностью используется гетеротрофными организмами и лимитирует их численность. Одновременно продукция растительности является важным мерилом трансформации солнечной энергии биотой и обмена веществ между биотой и окружающей абиотической средой.
Продукционный процесс в геофизическом аспекте
Цепь сложных превращений веществ и энергии, в результате которых фотосинтезирующие растения создают органическое вещество из неорганических соединений, называют продукционным процессом. Он включает в себя фотосинтез, дыхание и транспирацию растений. Как весь продукционный процесс, так и составляющие его частные процессы развиваются в природных комплексах в тесной зависимости от среды, прежде всего от солнечной радиации гидротермического режима.
Фотосинтез. В основе продукционного процесса лежит фотохимическое преобразование энергии, при котором фотосинтетически активная радиация (ФАР) поглощается и превращается в энергию химических соединений органического вещества. Это преобразование происходит в хлоропластах посредством хлорофилла. Роль главного фотосинтезирующего органа растения выполняет лист. Листья дают 60 — 95% суммарной продукции фотосинтеза растительного покрова, остальное приходится на долю стеблей, ветвей и других зеленых органов (Росс, 1975).
Процесс фотосинтеза в общем виде выражается уравнением
СО2+2Н2О ФАР/хлорофилл---(НСОН)+Н20+02,
из которого следует, что между поглощением СО2 и количеством образуемого органического вещества (НСОН) существует прямая зависимость. Участие азота и ряда других элементов минерального питания в фотосинтезе не оказывает влияния на эту зависимость.
Валовой фотосинтез и дыхание растений. Чистая продукция. Во время вегетации параллельно развиваются процессы фотосинтеза и дыхания зеленых растений. Их интенсивность зависит от температуры растений, но максимумы проявления не совпадают. Разность между валовым фотосинтезом (брутто-фотосинтез) и дыханием
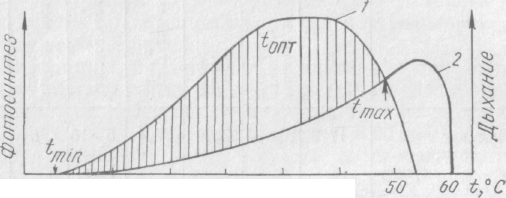
Рис. 8. Зависимость фотосинтеза (/) и дыхания (2) от
температуры (по Лархеру, 1978):
заштрихованная область — 'нетто-фотосинтез; ^rnin— температурный
минимум; <оп — температурный оптимум; /тах—температурный
максимум
определяет уровень чистой продукции (нетто-фотосинтез) (рис. 8).
Связь между чистой продукцией (Р), валовой ее величиной (Рв) и затратами на дыхание (D) можно выразить уравнением Р = РВ—D.
Транспирация. Диффузия водяного пара и диоксида углерода осуществляется через устьичный аппарат, поэтому транспирация и усвоение углерода в процессе фотосинтеза тесно связаны друг с другом. При этом градиент концентрации диоксида углерода между наружным воздухом и хлоропластами гораздо меньше градиента упругости водяного пара между внутренней средой листа и окружающим воздухом, если последний не насыщен влагой. В связи с этим испарение воды путем транспирации идет гораздо интенсивнее, чем поглощение диоксида углерода. Даже при закрытых устьицах, когда поглощение его прекращается, вода через кутикулу продолжает испаряться. В целом растение вынуждено приспосабливаться к жажде и голоду.
В то же время транспирация способствует поступлению в растение через корневую систему с водными растворами элементов минерального питания, необходимых для синтеза органического вещества. Правда, потери воды на транспирацию нередко чрезмерны. В среднем на 1 г сухого вещества чистой продукции в умеренных широтах травянистые растения расходуют на транспирацию 520—700, деревья — часто 170—340 г воды, примерно такие же показатели транспирации у высокопродуктивных растений. Однако для условий влажного тропического леса характерны иные показатели затрат транспирированной воды па единицу образованного органического вещества. Здесь воздух нередко насыщен влагой, что снижает расход транспирированной воды на единицу чистой продукции в несколько раз по сравнению с сообществами умеренных широт, где в течение вегетационного периода относительная влажность воздуха часто поддерживается на уровне 50—75%. Вместе с тем в любых природных условиях при низком содержании элементов минерального питания в почве расход воды па транспирацию возрастает, а на плодородных почвах снижается. Количественные соотношения образованного органического вещества и расхода транспирированной воды позволяют выявить своеобразие энергетики продукционного процесса растительного сообщества. В частности, в широколиственно-хвойных лесах на западе Восточно-Европейской равнины при создании 1 г сухого органического вещества транспирируется примерно 350 г воды. Учитывая, что калорийность 1 г сухого органического вещества равна 19,7 кДж, получаем, что при фиксации в чистой продукции 1 единицы энергии на транспирацию расходуется 43,9 таких же единиц. Отсюда следует, что при ассимиляции в нетто-продукции 1,5% энергии радиационного баланса вегетационного периода на транспирацию расходуется 65,85, на дыхание растений — около 1 и на теплообмен с атмосферой — примерно 31%. КПД транспирации (отношение энергии, содержащейся в чистой продукции, к энергии, затраченной на транспирацию) в этом случае — 2,28. Для сравнения можно указать, что по данным Ю. Л. Раунера (1973), в условиях центральной лесостепи Среднерусской возвышенности данный показатель при температуре воздуха 20—30°С составляет 2—2,5. При радиационном балансе вегетационного периода, равном 150,7 кДж/см2, чистая продукция может достигать в среднем 11,5 т/га-год, что является вполне реальной величиной для широколиственно-хвойного леса па западе Восточно-Европейской равнины в относительно благоприятных местоположениях.
При рассмотрении энергетики продукционного процесса обычно учитываются поступающая к земной поверхности суммарная солнечная радиация, а также поглощенная радиация и ФАР, но, как правило, игнорируется радиационный баланс земной поверхности, который имеет прямое отношение к продукционному процессу, а отряженная радиация и эффективное излучение в нем не принимают участия.
Зависимость продуктивности растительных сообществ от факторов среды
Как весь продукционный процесс, так и формирование чистой продукции растительности (ниже именуемой продукцией растительности) происходит в тесной зависимости от абиотической среды. Создание растениями органического вещества в виде продукции в течение вегетационного периода контролируется в природном комплексе рядом факторов среды, среди которых первостепенное значение имеют: энерго- и влагообеспеченность, обеспеченность растений элементами минерального питания, концентрация диоксида углерода в приземном слое воздуха, экстремальные природные явления, а также своеобразие самих растительных сообществ.
Энергообеспеченность природного комплекса. Накопление органического вещества в продукции растительности сопровождается большими затратами энергии, причем, как было показано выше, преобладающая ее часть идет на транспирацию и теплообмен с атмосферой, а сравнительно небольшая — ассимилируется в продукции. Но следует помнить, что вся энергия радиационного баланса земной поверхности с растительным покровом (т. е. радиационного баланса природного комплекса) если не прямо (через продукционный процесс), то косвенно (через термический режим среды) в течение вегетационного периода оказывает влияние на формирование продукции растительности. Поэтому одним из важнейших показателей энергообеспеченности природного комплекса выступает его радиационный баланс (В), а не суммарная солнечная радиация (Q). Из уравнения радиационного баланса земной поверхности B = Q—R—/ видно, что отраженная радиация (R) и эффективное излучение (/) не могут принимать участия в экзогенных физических процессах, включая продукционный процесс. Причем отраженная радиация и эффективное излучение вместе в аридных зональных природных комплексах составляют около 50—70% (по мере нарастания аридности их совместная доля возрастает), а в гумидных—примерно 40—50% суммарной солнечной радиации.
ФАР и инфракрасная радиация практически в равной мере участвуют в формировании радиационного баланса. Поскольку на долю ФАР приходится почти 1/2 суммарной радиации, а на транспирацию даже в наиболее продуктивных сообществах идет в 15—25 раз и больше энергии, чем ее фиксируется в продукции растительности, постольку ФАР в продукционном процессе расходуется преимущественно не на фотосинтез, а на транспирацию. Это вместе с тем свидетельствует об обеспеченности растительности ФАР в достаточной мере. Если листья нижних ярусов сообществ получают недостаточное количество ФАР, то в еще большей степени им не хватает энергии, необходимой для транспирации.
Основные энергозатраты радиационного баланса земной поверхности за вегетационный период (Bv) сводятся к затратам на испарение (LEv, где L — скрытая теплота испарения, Еv-—испарение) и турбулентный теплообмен с атмосферой (Тv). Расход энергии на фотосинтез (аРв, где а — калорийность сухого растительного вещества, Рв— валовая продукция растительности) не превышает нескольких процентов величины радиационного баланса, что обычно находится в пределах точности его вычисления. Поток тепла от подстилающей поверхности к почвогрунтам за вегетационный период (Av) в целом равен нулю, так как начало и конец этого периода фиксируются по одной и той же температуре воздуха. Но за часть вегетационного периода величина Av существенно отличается от нуля.
Тепловой баланс земной поверхности вегетационного периода по аналогии со средним годовым тепловым балансом (Будыко, 1977) с учетом расхода энергии на формирование валовой продукции растительности можно записать в виде уравнения
Bv = Lev +Tv + Av + aPb.
Величина LEV имеет непосредственное отношение к образованию продукции растительности, так как нередко преобладающую или по крайней мере значительную долю испарения (Ev) составляет транспирация. Часть Еv, включающая физическое испарение с поверхности растительного покрова и почвы, способствует повышению упругости водяного пара воздуха и тем самым косвенно благоприятствует уменьшению транспирации. Энергия теплообмена с атмосферой в большой степени определяет термический режим среды, прежде всего ее воздуха, что отражается на интенсивности фотосинтеза, дыхании растений и формировании нетто-продукции.
Отношение между основными составляющими теплового баланса земной поверхности за вегетационный период (Tv/LEv) в природных комплексах суши Земли изменяется в широких пределах. В гумидных условиях, где растительность обеспечена влагой в достаточной мере, Tv /LEv обычно менее единицы, в исключительных случаях снижается до нуля и даже становится отрицательной величиной. В частности, весьма небольшие значения этот показатель имеет во влажных тропических и субтропических лесах, а на болотах, в результате адвекции тепла с ближайших дренированных участков, может быть отрицательной величиной. Аналогичные явления наблюдаются в тугаях по долинам рек, протекающих через пустыни. Напротив, в аридных природных комплексах преобладающая часть радиационного баланса идет на теплообмен с атмосферой, поэтому Tv/LEv, здесь больше единицы, в пустынях может достигать б — 100 и более.
Надежными показателями энергообеспеченности являются суммы средних суточных температур воздуха выше 5 и 10°С. Между величиной радиационного баланса вегетационного периода и суммой температур выше некоторого предела при прочих равных условиях существуют определенные количественные соотношения. Они более заметно изменяются с изменением зональных природных условий и менее — в пределах природной зоны.
Величина отношения суммы температур выше 5 °С к радиационному балансу вегетационного периода более высоких значений достигает, как правило, в аридных зональных комплексах, где сравнительно высокие потери энергии на эффективное излучение несколько снижают величину радиационного баланса; одновременно значительная (или даже преобладающая) доля последней идет на теплообмен с атмосферой, что обеспечивает высокую температуру воздуха в течение вегетационного периода. Напротив, в гумидных комплексах этот показатель в связи с особенностями энергетического баланса несколько ниже. В то же время он возрастает от полярных широт к экваториальным как в гумидных (примерно от 13 в лесотундрах до 28°С/кДж/см2 во влажных тропических лесах), так и в аридных (от 16 в степях умеренных широт до 34 °С/кДж/см2 и более в тропических пустынях) условиях.
Таким образом, достаточно надежными показателями энергообеспеченности природного комплекса и его продукционного процесса могут быть радиационный баланс земной поверхности за вегетационный период и сумма температур воздуха этого периода.
Влагообеспеченность природного комплекса. Влагообеспеченность наряду с энергообеспеченностью играет важную роль в продукционном процессе и тем самым в образовании продукции растительности в природном комплексе. Как недостаток, так и избыток влаги отрицательно отражаются на фотосинтезе и продуктивности растительности.
Нередко за показатель влагообеспеченности природного комплекса принимается годовая сумма осадков, но при этом не учитывается, что часть осадков идет на сток, покидает комплекс и в продукционном процессе участия не принимает (исключение составляют резко аридные территории). Более того, со стоком выносятся элементы минерального питания растений, что приводит к снижению продуктивности растительности. Поэтому количеством атмосферных осадков за год не является достаточно точным показателем влагообеспеченности, особенно в гумидных условиях, где сток достигает значительной величины.
В соответствии с уравнением водного баланса территории за многолетний период годовая сумма осадков (S) идет на формирование стока (U), который имеет поверхностную и подземную составляющие, и суммарного испарения (Е) (Мировой водный баланс и водные ресурсы Земли, 1974; Львович, 1974), т. е. S=U + E.
Соотношение между составляющими водного баланса комплекса устанавливается в зависимости от его энергообеспеченности и особенностей теплового баланса, что наглядно проявляется на примере зональных комплексов. В резко аридных комплексах, в частности в пустынях умеренных, субтропических и тропических широт, практически все осадки идут на испарение, сток отсутствует, преобладающая часть радиационного баланса расходуется на теплообмен с атмосферой. Во влажных тропических лесах, где нередко почти весь радиационный баланс расходуется на испарение, последнее часто составляет менее 1/2 годовой суммы осадков и сток может превышать величину испарения.
Поскольку часть осадков со стоком выносится за пределы природного комплекса, его влагообсспеченность определяется той долей осадков, которая задерживается комплексом и используется на испарение. Влагообеспеченность наземных природных комплексов может достигать величины испаряемости, при этом часть влаги поступает в пределы комплекса иногда поверхностным или подземным путем извне. Именно так нередко происходит на поймах и в дельтах рек, понижениях рельефа. В то же время влагообеспеченность может падать почти до нуля в тропических пустынях, где единственным источником влаги являются осадки, годовая сумма которых может составлять 1—3 мм, например в оазисе Дахла и Асуане в Египте (Физико-географический атлас мира, 1964).
Влагообеспеченность природного комплекса, включая растительность, по величине соответствует суммарному испарению, которое, по замечанию Р. Слейчера (1970), из всех составляющих водного баланса наиболее тесно связано с физиологическими процессами в растении.
Составными частями суммарного испарения вегетационного периода (Ev) являются транспирация (Et),
физическое испарение с поверхности почвы (Еф) и испарение части атмосферных осадков, задержанных растительным покровом (Ер). т, е. Еv = Ет + Еф +Ер
В целом растительными сообществами при образовании одной единицы массы сухого вещества транспирируется от нескольких сот единиц массы в экстрагумидных комплексах (по влажных тропических лесах — 125) до 1000 — 1500 в экстрааридных местообитаниях (Молчанов, 1968, с. 91; Вальтер, 1968, с. 509; Рябчиков, 1968, с. 44; Будыко, 1977, с. 158). Нередко соотношение этих величин в умеренных широтах колеблется в пределах 1:300 — 1:700. У таких культурных растений, как рожь, овес, пшеница, ячмень, оно соответственно равно 1:630, 1 : 580, 1 : 540, 1 : 520, а, например, у бука потребность в транспирированной воде снижается до 170 г на 1 г сухого органического вещества, у дуба она в 2 раза больше. В то же время в каждом типе природного комплекса существует строго определенное соотношение между суммарным испарением и транспирацией. В зависимости от комплекса транспирация составляет 20 -80% суммарного испарения (Алпатьев, 1969, с. 184), увеличиваясь в общем от экстрагумидных комплексов к переходным и далее к аридным. Транспирация, выраженная в процентах от суммарного испарения, до минимума снижается во влажных тропических лесах, а максимума достигает в пустынях тропических и субтропических широт. В 30-летней дубраве, по данным А. А. Молчанова (1968, с. 16), на транспирацию приходится не менее 75% суммарного испарения вегетационного периода, а на долю испарения с поверхности почвы и травяного покрова падает не более 25%. Вероятно, с поверхности почвы в течение вегетационного периода при достаточно развитом древесном сообществе испарение составляет менее 10% суммарной его величины.
Атмосферные осадки, перехваченные растительным покровом, в основном подвергаются испарению. Чем больше индекс листовой поверхности, тем больше жидких осадков при прочих равных условиях задерживается сообществом и соответственно больше испаряется влаги. В умеренных широтах на кронах сосняков свыше 15-летнею и дубрав свыше 30-летнего возраста задерживается до 20 — 30% жидких осадков, при этом испарение с кроны 30-летней дубравы летом составляет более 13% годовой его величины (Молчанов, 1968). Под развитой травяной покров не проникает около 15% жидких осадков. Осадки задерживаются также мхом.
Испарение влаги с поверхности почвы и растительного покрова способствует увеличению содержания влаги в воздухе и росту его относительной влажности, что косвенно благоприятствует уменьшению транспирации. Таким образом, если транспирация является составной частью продукционного процесса, то физическое испарение с поверхности почвы и растительного покрова оказывает опосредованное положительное воздействие на продукционный процесс. Следовательно, суммарное испарение вегетационного периода прямо или косвенно связано с формированием продукции растительности.
Обеспеченность растений элементами минерального питания. В растениях обнаружена почти половина химических элементов периодической системы Менделеева. Не исключено, что в них содержатся практически все известные на Земле элементы в крайне ничтожных количествах, для установления которых современные аналитические приемы еще недостаточно точны.
Химические элементы, кроме углерода и отчасти свободного (несвязанного) кислорода, поступают в растения через корневую систему в процессе минерального питания с водными растворами.
Это прежде всего макроэлементы минерального питания (N, Р, К, Са). Их содержание в живых организмах, в частности в растениях, исчисляется десятыми и сотыми долями процента от сырой массы. Каждый из них оказывает определенное влияние на фотосинтез. Например, в процессе фотосинтеза образуются органические вещества — углеводы, аминокислоты и белки. Азотистые продукты фотосинтеза, т. е. аминокислоты и белки, составляют среди продуктов ассимиляции диоксида углерода около 25—35%, причем на долю белков приходится 1,5—4%. «По-видимому, лист является основным местом синтеза аминокислот и белка в зеленом растении» (Андреева, 1969, с. 160). Обеспечение растений минеральным азотом в значительной мере определяет интенсивность фотосинтеза.
Концентрация микроэлементов (В, F, Mg, Fe, Co, Си, Zn, Se, Вг, Mo, I и др.) в организмах ничтожна, но тем не менее они воздействуют на физиологические процессы в растениях, влияя на ход фотосинтеза и формирование продукции растительности (Молчанов, 1968; Школьник, 1970; Ягодин, 1970).
В организмах имеются также ультрамикроэлементы (например, Pb, Se, Cd), содержание которых не превышает п-10~4%. Их роль пока неизвестна (Чсртко, 1981). Элементы минерального питания должны быть сбалансированы между собой и относительно ресурсов тепла и влаги. Только в этом случае они оказывают оптимальное воздействие на количество и качество продукции растительности. Их несбалансированность может вызвать существенное снижение величины продукции. При благоприятных условиях обеспечения ФАР, теплом и влагой, но недостаточном минеральном питании уровень продукции растительности определяется минеральным питанием.
Растительные сообщества редко оптимально обеспечены минеральным питанием в соответствии с их ресурсами тепла и влаги. В то же время содержание элементов минерального питания в почве заметно изменяется при переходе от одного природного комплекса к другому. В средней полосе России наибольшим запасом доступных для растений элементов питания (и следовательно, наибольшим плодородием) отличаются дренированные среднесуглинистые почвы. Легкие по механическому составу почвы обладают незначительным содержанием питательных веществ, но хорошей воздухопроницаемостью. Напротив, тяжелые почвы содержат большое количество элементов минерального питания, но вследствие слабой воздухопроницаемости и обычно плотного сложения корневая система растений не может в полной мере их усваивать, что снижает плодородие этих почв.
В дренированных местообитаниях, где влажность почвы в течение вегетационного периода не превышает величины наименьшей влагоемкости, в почвенные поры поступает воздух, а вместе с ним кислород в количестве, обеспечивающем нормальное развитие корневой системы и растения в целом. Однако при избыточном увлажнении почвы и замедленном оттоке почвенных вод условия жизни растений ухудшаются из-за слабого газообмена между почвой и атмосферой. При этом корни растений недостаточно снабжаются кислородом, в почве накапливаются недоокисленные органические соединения, а в почвенном воздухе — вредные для растений газы (метан, сероводород, диоксид углерода). Все это отрицательно сказывается на развитии растений и их продуктивности. Однако в отдельных местообитаниях с избыточным увлажнением почвы растения развиваются вполне удовлетворительно. Это наблюдается там, где почвенные воды, постоянно обновляясь, омывают корневую систему растений, доставляют ей необходимые элементы минерального питания и кислород, одновременно удаляя различные вредные соединения. Такие сообщества можно встретить на поймах рек, а также иногда на пологих склонах, примыкающих к озерным котловинам или низинным болотам.
Различия в условиях плодородия и дренированности почв сказываются на продуктивности растительности низших по рангу комплексов (фации, урочища). В то же время при оценке средней продуктивности растительности зональных комплексов локальные особенности почв сглаживаются. Каждый зональный тип почвы, включая множество разновидностей, обладает определенными средними показателями плодородия и дренированности, что обусловлено совокупностью природных особенностей и прежде всего климатом.
Концентрация диоксида углерода в приземном слое воздуха. Фотосинтез возможен при определенной концентрации СО2 в воздухе. Высокопродуктивные растения при ярком освещении могут извлекать из воздуха СО2 до тех пор, пока его содержание не снизится до 20 частей на миллион или даже меньше. У остальных растений поглощение СО2 прекращается при уменьшении его содержания до 50- -80 частей на миллион (Лархер, 1978, с. 44).
Экспериментально установлено, что концентрация СО2 и интенсивность света примерно в одинаковой мере влияют на валовой фотосинтез: при высоких значениях обоих факторов достигается наибольшая его интенсивность, чем выше уровень одного фактора, тем больше реакция на фотосинтез другого. Иными словами, при достаточно высокой интенсивности света увеличение концентрации СО2 по крайней мере в несколько раз по сравнению с ее уровнем в атмосфере сопровождается ростом валового фотосинтеза, т. е. концентрация СО2 лимитирует интенсивность фотосинтеза (Хит, 1972).Однако в реальных условиях развития растительных сообществ концентрация СО2 изменяется в ограниченных пределах.
Использование огромного количества горючих полезных ископаемых как топлива в последние десятилетия привело к увеличению содержания СО2 в атмосфере. Если на рубеже 60—70-х гг. XX столетия средняя концентрация его в атмосфере составляла около 0,032%, или 320 млн.' по объему, то для 1975 г. приводится уже величина, равная 335 млн.-' (Лархер, 1978, с. 139). К середине 80-х гг. содержание СО2 в атмосфере, согласно прогнозу М. Е. Ляхова (1973), приблизится к 340 млн.
Концентрация СО2 в приземном слое воздуха обычно выше, чем в среднем в атмосфере. Одновременно она колеблется в известных пределах в зависимости от местных условий — типа природного комплекса и фитоценоза, характера циркуляции воздушных масс, времени года и суток, характера погоды.
Летом в лесах средней полосы европейской территории СССР в приземном слое воздуха наибольшая концентрация СО2 отмечена в относительно более продуктивных березовых и ясеневых, заметно меньшая — в кленовых, а также в сосновых древостоях. Если принять содержание СО2 на открытом месте у поверхности почвы в середине дня за 100%, то в типе леса сосняк-беломошник оно будет 106, в кисличнике— 122, долгомошнике— 106 и в сфагновом сосняке—102%. Причем в густых древостоях этот показатель может достигать 148% по сравнению с открытым местом (Молчанов, 1968, с. 64).
На протяжении вегетационного периода содержание СО2 в приземном слое воздуха на суше подвержено колебаниям в течение суток. Ночью в результате дыхания самих растений и микроорганизмов почвы концентрация СО2 у поверхности почвы в лесу превосходит ее средний показатель в атмосфере на 25% и более, а днем, особенно на уровне наибольшего расположения ассимилирующих органов, падает ниже среднего показателя. Концентрация СО2 изменяется, хотя и менее заметно, по сезонам года, если последние достаточно определенно выражены. Так, к северу от 30° с. ш. содержание СО2 в приземном слое воздуха возрастает за зимний период на 8— 9 млн.~' по объему от среднего показателя, достигая максимума в апреле, но к концу вегетационного периода (в сентябре) становится на 10—11 млн.-1 ниже среднего содержания в атмосфере.
Установлено, что концентрация СО2 над Мировым океаном в полярных широтах несколько больше, чем в тропических. Различия, вероятно, не выходят в среднем за пределы 10—20 млн.-1 Океан выступает как естественный регулятор содержания СО2 в атмосфере. При уменьшении парциального давления диоксида углерода в воздушной среде океан выделяет его, при увеличении— поглощает. В то же время, по мнению А. П. Виноградова (1967), океан, действуя как гигантский насос, поглощает СО2 из воздуха в холодных областях и выделяет в атмосферу — в тропических.
В целом в элементарных природных комплексах содержание СО2 в приземном слое воздуха в течение вегетационного периода колеблется в ограниченных пределах и сравнительно слабо изменяется в региональных комплексах при следовании из полярных широт в тропические. Поэтому можно считать, что при прочих равных условиях концентрация СО2 в атмосфере повсеместно определяет примерно одинаковый уровень первичного продуцирования органического вещества. В то же время комплексы всех уровней различаются по интенсивности метаболизма СО2, который наиболее высок там, где продукция растительности достигает наибольших значений (влажные тропические и субтропические леса, высокопродуктивные агросистемы), и существенно снижается при крайне небольшой продуктивности (арктические тундры, бесплодные пустыни, ледники).
В коренных природных комплексах, где все компоненты оптимально сбалансированы между собой, практически вся продукция растительности подвергается деструкции в результате дыхания гетеротрофных организмов. Это значит, что в среднем в течение года при образовании продукции растительности потребляется примерно столько же диоксида углерода, сколько его освобождается в результате разложения мертвых растительных остатков подстилки и почвы («дыхание почвы»). В целом из биотического круговорота выводится и подвергается захоронению ежегодно около 0,8%, углерода, используемого при формировании продукции растительности. Эта убыль восполняется без вмешательства человека в результате выделения СО2 из под океана и при вулканических извержениях,
Влияние экстремальных природных явлений на продуктивность растительности. Факторы, влияющие на формирование продукции растительности, не остаются строю постоянными. Обычно они изменяются от года к году, что приводит к существенным колебаниям годичной продукции. Уменьшение величины продукции растительности относительно ее среднего многолетнего уровня вызывает ряд природных явлений. К ним относятся: засухи и суховеи; избыточное выпадение атмосферных осадков, обычно сопровождающееся снижением ресурсов тепла; заморозки в течение вегетационного периода; градобития, ливни и ураганы, повреждающие фотосинтезирующие органы растений; водная и ветровая эрозия, уменьшающая плодородие почвы. Кроме того, запыленность атмосферы в результате вулканических извержений снижает величину поступающей к земной поверхности суммарной солнечной радиации и энергообеспеченность природных комплексов.
Вместе с тем за многолетний период примерно в 50% случаев годичная продукция при прочих равных условиях оказывается выше средней нормы. Это имеет место в те годы, когда в течение вегетационного периода высокие показатели энергообеспеченности (включая ресурсы тепла) сочетаются с достаточным увлажнением, а неблагоприятные природные явления отсутствуют.
Исходя из величины колебаний ширины годичного кольца у деревьев за отдельные годы (Молчанов, 1970; Долгушин, 1973), можно заключить, что годичная продукция лесов умеренных широт (Карельский перешеек, средне- и северотаежные районы Приобья Западной Сибири, Окско-Донская равнина) может изменяться за многолетний период примерно на один порядок под влиянием погодно-климатических условий. По этой же причине урожайность культурных растений подвержена колебаниям. В частности, урожайность зерновых культур на европейской территории России по десятилетиям снижалась в неурожайные засушливые годы: до 71,5% относительно наиболее урожайного года — в 60-е гг. и почти до 62% — в 70-е. Различия в урожайности по годам еще более резко проявляются на ограниченных площадях в аридных районах (южная степь, полупустыня). При общем росте урожайности зерновых культур абсолютный уровень потерь из-за погодно-климатических аномалий «в настоящее время имеет тенденцию не сокращаться, а возрастать» (Раунер, 1981, с. 146—147).
Влияние на годичную величину продукции своеобразия растительных сообществ. Каждое коренное растительное сообщество оптимально приспособлено к среде. Поскольку абиотическая среда на ограниченной территории нередко существенно изменяется, постольку одновременно изменяются сообщества и их годичная продукция. Последняя в двух ближайших сообществах может различаться по величине в несколько раз (например, сосняк-беломошник на дерново-подзолистой песчаной почве и дубрава кустарниковая на серой среднесуглинистой почве). Эти различия определяются особенностями плодородия почв, их влагообеспеченностью и характером растительных сообществ, неразрывно связанных со средой. Причем в коренных природных комплексах практически невозможно разграничить влияние абиотической среды и влияние самого растительного сообщества на величину его продукции
Однако при средней оценке продуктивности растительности зональных сообществ их локальные особенности нивелируются. В результате годичная величина продукции плавно изменяется как при переходе от одной природной зоны к другой, так и внутри зон. Эти изменения находятся в полном соответствии с такими показателями, как индекс листовой поверхности, содержание хлорофилла в листьях, продолжительность вегетационного периода. Что касается плодородия почвы, то оно, будучи связано с климатом, изменяется в целом вместе с зональными типами растительных сообществ. В то же время годичная величина продукции растительности резко различается в типичных зональных сообществах, развивающихся в экстремальных условиях среды, например, в арктических и тропических пустынях, с одной стороны, во влажных тропических лесах — с другой.
Соотношение продуктивности растительности и показателей абиотической среды
Активное развитие отдельных растений и растительных сообществ происходит на протяжении вегетационного периода, когда формируется продукция растительности. Продолжительность этого периода в арктических пустынях определяется временем проявления положительной средней суточной температуры. Но уже на юге тундровой зоны и южнее вегетационный период выделяется как период с устойчивой средней суточной температурой выше 5 °С. В высоких и умеренных широтах он длится меньше года, а в низких (субтропических и тропических) — весь год. При этом в аридных природных комплексах вегетация растений наряду с термическим фактором лимитируется ресурсами влаги.
Средняя годовая величина продукции растительности за многолетний период (Р) выступает как функция ряда геофизических переменных среды: ресурсов ФАР и тепла, находящих одновременно свое выражение в радиационном балансе земной поверхности вегетационного периода (Вv); ресурсов влаги, реализуемых на суммарное испарение и потому равных величине испарения вегетационного периода (Ev); обеспеченности почвы доступными элементами минерального питания (т); концентрации диоксида углерода в атмосфере. Поскольку содержание диоксида углерода в воздухе колеблется в ограниченных пределах и синхронно в целых географических поясах, этот показатель на достаточно обширных территориях действует одинаково, представляя собой своеобразный фон, в связи с чем его можно исключить из числа переменных величин. Тогда зависимость средней годовой величины продукции растительности от геофизических параметров среды выразится в следующем виде:
P = f (Bv, Еv, т).
При этом допускается, что стихийные природные явления в течение многолетнего периода имеют некоторую постоянную ритмичность и потому не оказывают влияния на среднюю величину продукции.
Соотношение годичной продукции растительности и радиационного баланса земной поверхности вегетационного периода (Вv). Показатель Bv в низких широтах соответствует годовому радиационному балансу. В умеренных широтах (например, на территории России) он примерно на 10% меньше, а в арктических пустынях составляет около 80%. годовой величины. Так, радиационный баланс земной поверхности вегетационного периода в арктических пустынях, в местах, свободных в теплое время года от снега и льда, равен нескольким десяткам килоджоулей на квадратный сантиметр. По мере продвижения к экватору в зональных природных комплексах на равнинах его средняя величина возрастает, достигая во влажных тропических лесах 300—380 кДж/см2.
Вместе с тем она может изменяться на относительно небольшой территории в некоторых пределах в зависимости от подстилающей поверхности, в первую очередь от свойств растительного покрова и степени увлажнения почвы. В частности, в пустынях умеренных, субтропических и тропических широт величина Bv на поймах рек, где формируется пышная древесно-кустарниковая растительность на достаточно увлажненной аллювиальной почве, может на 50% и более быть выше, чем па междуречьях с крайне разреженной растительностью и искусственной почвой.
Среднее значение интенсивности радиационного баланса земной поверхности за вегетационный период (Bv/tv, где tv— продолжительность вегетационного периода) у зональных природных комплексов от тундровой зоны до влажных тропических лесов колеблется от 0,70 до 1,00 кДж/см2-сут. Лишь в исключительных случаях оно выходит за эти пределы. Средняя интенсивность Вv практически почти одинакова по всех природных зонах суши Земли.
Сравнение величин Вv на территории России и других стран субтропических и тропических широт с показателями средней продуктивности растительности суши Земли картосхем Н. И. Базилевич и Л. Е. Родина (1967; 1971) позволяет сделать следующее заключение. В гумидных зональных природных комплексах, где годовая сумма атмосферных осадков больше испаряемости, увеличение Вv сопровождается ростом годичной продукции растительности. Это отчетливо прослеживается па территории России при движении с севера на юг, от арктических пустынь к лесостепной зоне, а также при сравнении лесных комплексов умеренных, субтропических и тропических широт. В аридных зональных комплексах, особенно в пустынных и полупустынных их разновидностях, рост величины Вv, как правило, при одновременно уменьшающейся обеспеченности растений влагой вызывает снижение продукции растительности.
Следовательно, хотя продуктивность растительности, несомненно, зависит от радиационного баланса земной поверхности, эта зависимость по-разному проявляется в гумидных и аридных зональных комплексах. Вместе с тем такая составляющая теплового баланса, как затраты тепла па испарение за вегетационный период (LEV), а также доля радиационного баланса этого периода, идущая на испарение (LEv/Bv), изменяются в соответствии с величиной продукции (Зубов, 1978, с. 64— 66).
Соотношение годичной продукции растительности и ресурсов влаги. Ресурсы влаги, прямо или косвенно связанные с формированием продукции растительности, соответствуют испарению вегетационного периода (Ev). Величина Ev тесно связана с продукционным процессом и продуктивностью растительности.
Для определения испарения с достаточно больших по площади территорий суши используются различные методы. Нами принята методика определения испарения А. Р. Константинова (1968) —по температуре и влажности воздуха, измеренным на метеостанциях, с внесением соответствующих поправок. А. Р. Константинов предложил графики для расчета испарения за сутки и год. Преимущество указанной методики заключается в том, что она позволяет сравнительно просто получать данные по испарению за требуемый промежуток времени, например за вегетационный период.
Как отмечает автор методики, в ряде природных комплексов репрезентативность показаний метеостанций нарушается, что влечет завышение показателей испарения. Это прежде всего относится к аридным комплексам (пустыни, полупустыни, в меньшей мере степи), где метеостанции расположены среди хорошо обводненных оазисов, и районам на побережьях водоемов (морей, озер), которые оказывают влияние на температуру и влажность воздуха суши. Поэтому показатели испарения, рассчитанные по методике А. Р. Константинова, для аридных и прибрежных районов необходимо корректировать с величинами испарения, установленными исходя из уравнения водного баланса.
Испарение вегетационного периода в низких широтах соответствует годовому испарению, в умеренных широтах оно снижается относительно годовой величины на 10—25%, а в тундрах и арктических пустынях—еще больше.
По существу, испарение является совокупным показателем ресурсов влаги и энергии продукционного процесса. В гумидных природных комплексах, где значительная часть атмосферных осадков (примерно 20— 70% годовой суммы и более) идет на формирование стока, испарение в течение вегетационного периода определяется в основном ресурсами тепла. При этом затраты тепла на испарение в типичных зональных природных комплексах не превышают величину радиационного баланса, так как часть его используется для теплообмена с атмосферой. В аридных природных комплексах испарение обычно лимитируется количеством выпадающих осадков. В частности, во всех типах пустынь земного шара атмосферные осадки практически полностью расходуются на испарение. Более того, в тех случаях, когда корневая система достигает капиллярной каймы, испарение в аридных условиях может во много раз превышать годовую сумму осадков, что наблюдается, например, на поймах и в дельтах рек. Величина испарения определяется здесь ресурсами тепла с учетом его адвекции с соседних участков.
На ограниченной территории величина Ev в значительной мере зависит от мезо- и микрорельефа, характера почвогрунтов и растительности. Мезо- и микропонижения нередко обладают более тяжелыми по механическому составу почвогрунтами, чем ближайшие поднятия. Они лучше обеспечены влагой и отличаются в дренированных условиях относительно высоким испарением. Здесь обычно, за исключением резко аридных комплексов, располагаются и наиболее продуктивные растительные сообщества, развивающиеся в сравнительно благоприятных условиях влагообеспеченности.
Величина испарения вегетационного периода суши, где формируется растительный покров, изменяется от нескольких десятков миллиметров в арктических пустынях и в наиболее засушливых районах тропических пустынь до 1200 мм и выше—во влажных тропических лесах. Средняя годичная продукция растительности в арктических и тропических пустынях составляет менее 1 т/га, а во влажных тропических лесах достигает 30 т/га и более.
Таким образом, между величиной испарения вегетационного периода и продукцией растительности существует прямая связь.
Зависимость годичной продукции растительности от совместного влияния радиационного баланса земной поверхности и испарения вегетационного периода. Средняя годичная продукция растительности (Р) достаточно крупного региона тесно связана с радиационным балансом земной поверхности вегетационного периода (Вv) и испарением того же периода (Ev). Эта зависимость была рассмотрена ранее (Зубов, 1977; 1978). Она выражается посредством формулы
(1)
Р = 0,42 LE2v/Bv, т/га-год.
На продукцию растительности комплексов топологического уровня (фация, урочище) заметное влияние оказывают локальные особенности почвогрунтов. При определении продукции этих комплексов в формулу (1) необходимо вводить коэффициент обеспеченности растений доступными элементами минерального питания, т. с. относительного плодородия почвогрунтов (т). Тогда эта формула примет вид
(2)
Р = 0,42т LEv /Bv, т/га-год.
Значение коэффициента т может изменяться в зависимости от свойств почвогрунтов от нуля на выходах на поверхность скальных пород, лишенных почвы, до двух и выше —на почвах, наиболее обеспеченных доступными для растении элементами минерального питания.
Среднюю годичную продукцию растительности достаточно крупного комплекса (или региона) можно определить также с помощью графика (рис. 9).
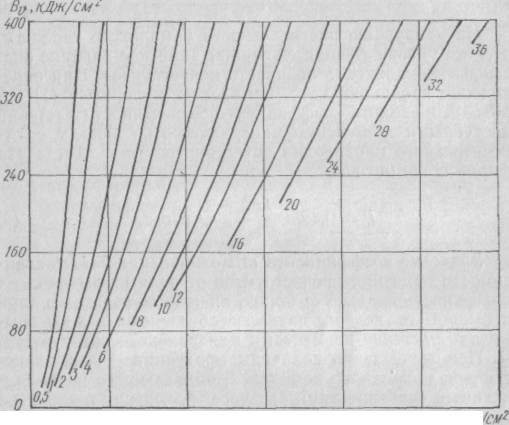
20 40 60
Рис. 9. График определения средней продуктивности растительности (т/га-год) по данным радиационного баланса (Bv) и испарения (Ev) за вегетационный период
В дренированных местоположениях при расчете средней годичной продукции растительности затраты энергии на испарение за вегетационный период (LEV) не могут превышать в тропических и субтропических широтах 0,75 Вv, в более высоких широтах они заметно меньше, а в тундре снижаются примерно до 0,5 Вv и ниже. Следует особо отметить, что на болотах, где вследствие адвекции тепла величина LEV может быть равна Вv или быть даже больше Вv при расчете средtгодичной продукции региона необходимо ограничивать значение Ev таким образом, чтобы 2 LEv/BV не превышало 0,75Однако по ряду районов обнаружены отклонения расчетных показателен от соответствующих значений картосхем—обычно не более ±10—15%, что находится в границах точности определения продукции на местности
В ряде случаев отклонения можно объяснить также не вполне точным измерением на местности, с одной стороны — продукции растительности, с другой — радиационного баланса и испарения вегетационного периода. Примеры, свидетельствующие о недостаточно точном отражении на картосхемах годичного прироста растительности, приводились ранее. Известные затруднения в некоторых природных комплексах вызывает определение испарения вегетационного периода. Это, в частности, относится к саксауловым и кустарниковым пустыням на пустынных песчаных почвах России, для которых реальная величина испарения вегетационного периода обусловлена не только атмосферными осадками, но и отчасти доступностью грунтовых вод для корневой системы растений.
В гумидных комплексах по мере увеличения площади болот, например в лесной зоне Западно-Сибирской равнины, величина испарения вегетационного периода возрастает, а продукция растительности уменьшается. В результате расчетная продукция оказывается больше реально установленной на местности на 10 — 20%.
Соотношение величины продукции растительности, потребления диоксида углерода и продуцирования кислорода. При формировании продукции растительности устанавливаются определенные количественные соотношения между образованием органического вещества продукции, потреблением диоксида углерода и выделением кислорода. Поскольку в состав продукции растительности суши, кроме углерода, кислорода и водорода, входят зольные элементы и азот, согласно О. П. Добродееву (1974) при создании одной единицы массы сухого вещества продукции потребляется 1,88 таких же единиц диоксида углерода и выделяется 1,38 аналогичных единиц кислорода.
Средний показатель продукции растительного покрова одновременно характеризует степень метаболизма природного комплекса.
Определение средней максимально возможной годичной продукции растительности. В пределах достаточно крупного по площади региона формируется множество растительных сообществ, а на сельскохозяйственных нолях — агроценозов. При оптимальной сбалансированности всех факторов среды, влияющих на продукцию, максимальная величина последней колеблется между отдельными сообществами в некотором интервале. Средний показатель продукции растительности этих сообществ есть средняя максимально возможная се величина.
В природных комплексах продукция растительности крайне редко приближается к максимальному значению, что обычно связано с недостаточной влагообеспеченностью и нехваткой элементов минерального питания. При этом приток солнечной энергии к земной поверхности па вегетационный период в любом природном комплексе остается практически постоянным. Очень высокие показатели продукции в ряде случаев в умеренных широтах отмечаются у сообществ пойм и дельт рек. Так, продукция тростника в наиболее благоприятных условиях в дельте Амударьи, по данным 3. Г. Ганецкой (1971, с. ПО), достигает 72— 132 т/га -год воздушно-сухого вещества. В целом для дельты Амударьи она принимается равной 30 — 50 т/га 'год (Базилевич, Родин, 1971). Продукция сообществ крупнотравья по долинам рек Сахалина доходит до 31,8 т/га -год сухого вещества (Степанова, Рассохина, 1981.Однако большинство зональных сообществ даже в самых благоприятных условиях среды обладают меньшей продуктивностью.
Средняя максимально возможная продукция региона формируется при оптимально сбалансированной обеспеченности солнечной радиацией, влагой и элементами минерального питания. Содержание диоксида углерода и ноздухе практически остается постоянным на протяжении вегетационного периода.
Важнейшим показателем, лимитирующим максимальную продукцию сообщества, является солнечная радиация, интегральное выражение которой — радиационный баланс увлажненной земной поверхности вегетационного периода, Он одновременно указывает на ресурсы ФАР и тепла конкретного сообщества. При этом состав и содержание элементов минерального питания в почве и обеспеченность влагой должны находиться в соответствии с потребностью растений в них при данной интенсивности радиационного баланса, а сложение почвы должно отвечать оптимальному газообмену между корневой системой растений и припомненным слоем воздуха. По мнению А. М. Ллпатьева (1969, с. 283—284), «в целом за вегетационный период потребность в воде фитоценоза можно безошибочно приравнять к испаряемости, вычисляя ее любым способом, наиболее точно отражающим местные природные, в том числе климатические условия». На определенных стадиях развития растений их потребность в воде может даже превышать величину испаряемости. «Ко времени наступления бутонизации — цветения у однолетних или максимального прироста у многолетних растений потребность в воде фитоценозов всегда оказывается больше испаряемости, что предполагает наличие адвекции ресурсов тепла с окружающих фитоценоз территорий» (Алпатьов, 1969, с. 284). Это, по-видимому, справедливо для небольших по площади фитоценозов. Регионы, охватывающие в умеренных и полярных широтах значительные территории (провинции, область), путем адвекции не получают тепло, так как в течение вегетационного периода в них довольно часто вторгаются морские относительно холодные воздушные массы из умеренных и полярных широт. На суше (в лесной, лесостепной и степной зонах) они прогреваются и превращаются в континентальный воздух, для чего необходима энергия. Вхождение теплого тропического воздуха морского и континентального происхождения, судя по его повторяемости (Борисов, 1970), не компенсирует этих потерь.
Если затраты тепла на среднее максимально возможное испарение достигают величины радиационного баланса увлажненной земной поверхности вегетационного периода , то LE0v/B0v становится равным единице.
В гумидных комплексах умеренных широт при движении с юга на север понижается температура воздуха и почвы, что приводит к уменьшению испарения вегетационного периода
Распределение годичной продукции растительности на суше Земли. Величина продукции растительности изменяется на суше в строгом соответствии с абиотической средой. В крайне неблагоприятных условиях арктических пустынь величина годичной продукции в местах, не освобождающихся в теплое время года от снега и льда, приближается к нулю. По направлению к югу она постепенно нарастает, достигая на юге лесной зоны и в лесостепи
Здесь значительные ресурсы энергии и влаги приближаются к оптимальной сбалансированности. Южнее лесостепной зоны величина годичной продукции снижается по мере усиления аридности и в солянковых пустынях на широте Аральского моря падает до 1,6 т/га и ниже. Но в пустынных сообществах, развивающихся на песчаных пустынных почвах и сероземах предгорий Средней Азии, она может возрастать до 3—8 т/га. Наиболее благоприятные условия для формирования продукции плакорных сообществ складываются во влажных субтропиках Закавказья, где ее величина колеблется от 15 до 30 т/га-год. Еще более значительны показатели продукции на поймах и в дельтах рек пустынной зоны.
Одновременно на территории России отмечается тенденция уменьшения продуктивности растительности с запада на восток в тундровой, лесотундровой и лесной природных зонах. Наиболее четко она выражается при сравнении растительных сообществ внутри отдельных зон Восточно-Европейской равнины, с одной стороны, Средней и Северо-Восточной Сибири и Дальнего Востока — с другой. Это находится в прямой связи с сокращением продолжительности вегетационного периода и уменьшением ресурсов тепла и влаги. В результате средняя годичная продукция уменьшается в границах каждой гумидной зоны в направлении с запада на восток на 25—50%. Подобное снижение продукции выявлено также в лесостепной и степной зонах в пределах Восточно-Европейской и Западно-Сибирской равнин.
В горах величина продукции изменяется с высотой в зависимости от гидротермических условии равнин подножий.
На склонах гор, граничащих с гумидными природными комплексами равнин подножий, с поднятием сокращается продолжительность вегетационного периода, уменьшаются ресурсы энергии и влаги за этот период, что приводит к уменьшению продукции растительности, которая в нивальной зоне падает до ничтожной величины. Так происходит на территории России в горах, лежащих севернее лесостепной зоны, а также на горных склонах, подножия которых граничат с влажными субтропиками Закавказья.
Иная ситуация обычно складывается в том случае, когда у подножий гор развиваются резко аридные природные комплексы (пустыни, полупустыни). В горах с высотой сокращается длительность вегетационного периода, причем в предгорьях — незначительно. Однако радиационный баланс и ресурсы влаги вегетационного периода возрастают вместе с увеличением количества атмосферных осадков до определенной высоты, выше которой они постепенно сокращаются. Это, в свою очередь, обусловливает увеличение продукции с поднятием также до некоторой высоты, но выше се величина постепенно снижается, падая до крайне низких значений близ снеговой линии. В то же время в изолированных котловинах и долинах, где количество осадков вверх по склонам не возрастает, величина продукции, как правило, с высотой постепенно сокращается.
Некоторые особенности распределения средней максимально возможной годичной продукции растительности (Р0) на суше Земли. Показатели Р0 на равнинах суши возрастают практически от нуля па льдах полярных пустынь до 65 т/га-год и более в тропическом поясе. С учетом горных природных комплексов, где в условиях достаточного увлажнения продуктивность растительности в целом несколько снижается по сравнению с равнинами подножий, среднее значение Р0 принимается равным в тропическом поясе 65, субтропических поясах — 55, умеренных — 25, полярных — 6 т/га • год.
Произведенный расчет показывает, что средняя максимально возможная годичная продукция растительного покрова суши Земли составляет примерно 560,3 -109 Т (45,92 т/га). Это превышает величину современной годичной продукции суши почти в 5 раз.
На территории России средняя максимально возможная годичная продукция увеличивается почти от нуля на льдах арктических островов и горных ледниках до 45— 60 т/га на равнинах и в предгорьях Закавказья и юга Средней Азии.
Определение резерва продуктивности растительности суши Земли. Зная среднюю максимально возможную продуктивность (Р0) и реальную ее. годичную величину (Р) для конкретного региона, легко определить резерв продуктивности. Величина резерва продуктивности возрастает от 1—2 т/га-год в арктических пустынях до 60 т/га • год и более в тропических и субтропических пустынях.
Вторичная биологическая продуктивность
Зеленые растения (продуценты) в процессе фотосинтеза создают первичную продукцию из неорганических соединений, трансформируя ФАР в потенциальную химическую энергию. Гетеротрофные организмы (консументы и редуценты), потребляя готовые органические вещества, в свою очередь, создают биомассу — вторичную продукцию.
Консументы питаются разнообразными органическими веществами и делятся на три группы: 1) растительноядные животные (фитофаги), пожирающие ткани продуцентов (значительную часть продукции растительности), а также растительные и животные паразиты зеленых растений; 2) хищники первого порядка (нередко сравнительно мелкие хищники), питающиеся фитофагами, паразиты, обитающие на фитофагах; 3) хищники второго порядка (обычно относительно крупные хищники), питающиеся хищниками первого порядка, и паразиты хищников первого порядка.
Редуценты питаются мертвыми растительными остатками (соответствующими части продукции растительности, поступающей в спад), трупами животных и экскрементами. Они осуществляют разложение органического вещества до образования устойчивых минеральных соединений. К ним относятся животные, принадлежащие к разнообразным систематическим группам, а также бактерии и грибы.
Одни и те же продуценты могут служить кормом различным растительноядным животным, а последние — пищей разным хищникам. В результате в биоте создается множество трофических (кормовых) цепей, которые переплетаются между собой. Среди них обычно выделяют два основных вида: цепи выедания и цепи разложения Цепи выедания начинаются продуцентами, создающими первичную продукцию из минеральных соединений абиотической среды, продолжаются растительноядными животными, потребляющими первичную продукцию и, в свою очередь, служащими пищей для относительно мелких хищников, которых пожирают более крупные хищники, и т. д. При этом паразиты в трансформации органического вещества проявляют себя либо кафитофаги, либо как хищники.
Цепи разложения начинаются мертвым органическим веществом и продолжаются разнообразными преимущественно микроскопическими организмами-редуцентами.
Вторичная продуктивность, или скорость продуцирования органического вещества гетеротрофами, на всех уровнях во много раз меньше первичной продуктивности. При создании одной единицы массы вторичной продукции консументами и редуцентами на каждом трофическом уровне потребляется в несколько раз больше органического вещества. В связи с этим вторичное продуцирование органического вещества, по существу, выступает как деструкция первичной биологической продукции. В трофических цепях биоты (или биоценоза) передача энергии с одного уровня на другой осуществляется с относительно низкой эффективностью. В среднем в цепях выедания от 10 до 20% энергии, связанной растительным сообществом в годичной продукции, может быть передано травоядным, 10—20% энергии, поступающей в сообщество травоядных, может быть перенесено на уровень хищников первого порядка и т. д.. Следовательно, на уровень хищников второго порядка может быть передано не более 0,1 — 0,2% энергии первичной продукции..
В цепях разложения первичная продукция потребляется редуцентами нередко не в меньшем объеме, чем в цепях выедания. Причем передача энергий от органических соединений, которые являются кормом микроорганизмов, происходит с большим эффектом (достигает 15—30%).
Первичная продукция распределяется примерно поровну между фитофагами и редуцентами. Фитофаги ассимилируют около 10% энергии корма, а 90% се с экскрементами поступает редуцентам. Продукция фитофагов, в которой ассимилируется вряд ли более 1% энергии первичной продукции, потребляется хищниками первого порядка. Последние усваивают также около 10% энергии корма, а остальные 90% ее идет редуцентам. На уровень хищников второго порядка может быть передано, вероятно, не более 0,02% энергии первичной продукции, но и она используется ими для образования биомассы также с низкой эффективностью. При этом в результате миграции организмов энергия может поступать в геосистему извне и выводиться из нее.
Все группы животных в целом в цепях выедания при ассимиляции 10—20% энергии корма получают не более 10% энергии первичной продукции, а свыше 90% ее в коренной геосистеме используют редуценты, подвергая минерализации опад, экскременты и трупы животных. Редуценты в геосистеме выступают как основные деструкторы органического вещества и главные трансформаторы химически связанной энергии первичной продукции.