


Рис. 217. Импульсация двух ганглиозных клеток сетчатки (а и б) и их концентрические рецептивные поля (РП).
Тормозные зоны рецептивных полей заштрихованы. Показаны реакции на включение (1 и 4) и выключение (2 и 3) света при стимуляции световым пятном центра РП (I и 3) и его периферии (2 и 4).
рецептивного поля обладает максимальной световой чувствительностью, а на краях его чувствительность снижается. При увеличении размера светового пятнышка, вспыхивающего в пределах центра рецептивного поля, величина ответа ганглиозной клетки увеличивается (пространственная суммация). Возбуждение двумя стимулами близко расположенных ганглиозных клеток приводит к их взаимному торможению: ответы каждой клетки делаются меньше, чем при одиночном раздражении. В основе этого эффекта, обеспечиваемого амакриновыми клетками сетчатки, лежит горизонтальное или боковое торможение. Рецептивные поля соседних ганглиозных клеток частично перекрываются, так что одни и те же рецепторы могут вносить вклад в генерацию ответов нескольких нейронов. Благодаря круглой форме рецептивные поля ганглиозных клеток сетчатки производят так называемое точечное описание сетчаточного изображения: оно отображается очень тонкой мозаикой, состоящей из точек — возбужденных нейронов.
Такой же характер имеет и картина возбуждения в нейронных слоях подкоркового зрительного центра — наружного коленчатого тела, куда приходят волокна из сетчатки. Рецептивные поля этих нейронов также круглые, но меньшего размера, чем в сетчатке. Фазический характер их ответов также выражен больше, чем в сетчатке. На уровне наружного коленчатого тела происходит взаимодействие афферентных сигналов, идущих из сетчатки, с эфферентными сигналами из коры, а также из слуховой и других сенсорных
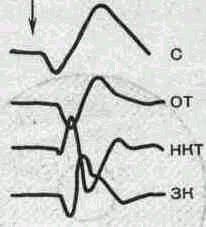
С — ВП сетчатки (ЭРГ); ОТ — ВП оптического тракта; НКТ — ВП наружного коленчатого тела и ЗК — ВП первичной проекционной области зрительной коры у наркотизированной кошки.
систем, опосредованными нейронами ретикулярной формации. Эти взаимодействия происходят в сложных синаптических структурах наружного коленчатого тела, куда, помимо перечисленных волокон, подходят также отростки клеток, осуществляющих торможение. Все эти взаимодействия обеспечивают дополнительную обработку зрительной информации — выделение в ней наиболее существенных компонентов и процессы избирательного зрительного внимания. Считается также, что в наружном коленчатом теле происходит важное для глубинного стереоскопического зрения послойное разделение сигналов, приходящих из правой и левой сетчаток.
Для оценки характера и направленности процессов переработки зрительных сигналов на разных уровнях анализатора используют регистрацию суммарных вызванных потенциалов, которые у животных можно одновременно исследовать во всех отделах системы, а у человека — в зрительной коре большого мозга (рис. 218). Сравнение вызванных ответов в сетчатке глаза на электроретинограмме и в коре позволяет во многих случаях провести точный дифференциальный диагноз и установить локализацию патологического процесса в зрительной системе человека.
Разряды нейронов наружного коленчатого тела по их аксонам поступают в затылочную часть коры головного мозга, где расположена первичная проекционная область зрения. Здесь у приматов и человека, происходит значительно более специализированная и сложная, чем в сетчатке и наружном коленчатом теле, переработка информации. Нейроны зрительной коры большого мозга имеют не круглые, а вытянутые (по горизонтали, вертикали или в одном из косых направлений) рецептивные поля небольшого размера. Благодаря этому они оказались способными осуществлять так называемые детекторные функции: выделять из всего изображения лишь отдельные его фрагменты той или иной ориентации и расположения и избирательно на них реагировать. Кроме того, разные клетки зрительной коры различаются по тонкости анализа изображений; одни из них, имеющие простые рецептивные поля, реагируют лишь на строго локальные и маленькие линии, другие (со сложными рецептивными полями) отвечают на такие изображения в большей, чем простые нейроны, части поля зрения.
В каждом небольшом участке зрительной коры по ее глубине сконцентрированы нейроны с одинаковой ориентацией и локализацией рецептивных полей в поле зрения. Они образуют колонку нейронов, проходящую вертикально через все слои коры. Колонка — пример функционального объединения корковых нейронов, осуществляющих сходную взаимодополняющую функцию.
Роль движения глаз для зрения
При рассматривании любых предметов важную роль играют движения глаз. Движения глаза осуществляются при помощи 6 мышц, прикрепленных к глазному яблоку несколько кпереди от его экватора. Это 2 косые и 4 прямые мышцы — наружная, внутренняя, верхняя и нижняя (рис. 219). Из них только наружная мышца поворачивает глаз прямо наружу, а внутренняя — прямо внутрь. Верхняя же и нижняя прямые мышцы поворачивают глаз не только вверх или вниз, но и немного внутрь, поэтому, чтобы повернуть глаз прямо вверх или вниз, к сокращению прямых мышц должно присоединяться сокращение косых мышц.
Движение глаз совершается одновременно и содружественно. Рассматривая более близкие предметы, необходимо сводить, а рассматривая более далекие предметы — раз-
Рис.
219.
Глазные мышцы.
1
—
верхняя косая;
2—
поднимающая веко;
3— верхняя
прямая;
4 —
наружная прямая;
5 —
нижняя прямая;
6—нижняя
косая;
7—зрительный
нерв.
Важная роль движений глаза в процессе зрения прежде всего определяется тем, что для непрерывного получения зрительной информации необходимо движение изображения по сетчатке. Как показали электрофизиологические исследования, импульсы в зрительном нерве возникают лишь в момент включения и выключения светового изображения. При непрерывном воздействии света на зрительные рецепторы импульсация в соответствующих волокнах зрительного нерва быстро прекращается и зрительное ощущение при неподвижных глазах и объектах исчезает. Если на роговице глаза укрепить маленькую присоску с источником светового раздражения, смещающимся вместе с глазом при его движении, так, чтобы раздражение действовало все время на одни и те же элементы сетчатки, то в этом случае человек будет видеть свет только в течение 1—2 с после его включения.
С помощью этого метода обнаружено, что глаз при рассматривании любого предмета производит не ощущаемые человеком непрерывные скачки. Вследствие этого изображение на сетчатке непрерывно смещается с одной точки на другую, раздражая тем самым новые фоторецепторы и вызывая вновь импульсацию в ганглиозных клетках и отходящих от них нервных волокнах. Продолжительность каждого отдельного скачка равна сотым долям секунды, а размер его не превышает 20°. Скорость скачка колеблется от 200° до 400° в секунду. Длительность интервалов между отдельными скачками, т. е. продолжительность фиксации взора на рассматриваемом предмете или светящейся точке, составляет в среднем 0,2—0,5 с, но может быть и больше. Чем сложнее объект, который рассматривает глаз, тем сложнее кривая его движения. Глаз человека как бы ощупывает контуры изображения, задерживаясь и возвращаясь к тем участкам, которые по тем или иным причинам привлекают особое внимание. Таким образом, человек получает более подробную информацию о деталях изображения.
Кроме скачков, глаз непрерывно мелко дрожит и дрейфует (медленно смещается с точки фиксации взора). Эти движения также играют роль в дезадаптации зрительных нейронов.