

6. Инициация трансляции.
Инициация в процессе синтеза белка означает не просто начало элонгации. Прежде всего, так как начало кодирующей последовательности мРНК не совпадает с началом полинуклеотидной цепи, а всегда находится, отступая от ее 5'-конца (иногда на значительное расстояние), необходимо точное узнавание первого кодона на внутренней части цепи. Это узнавание определяет не только начало полипептидной цепи, которая синтезируется, но и задает фазу всего дальнейшего считывания мРНК по триплетам, т. е. Абсолютно критично для всей аминокислотной последовательности полипептида. Другими словами, именно инициация определяет фиксированную точку на матричном полинуклеотиде, с которой начинается отсчет триплетов без запятых.
С другой стороны, инициация является главной точкой приложения механизмов регуляции синтеза белка на уровне трансляции. Регуляция на уровне трансляции у прокариот практически целиком сводится к разрешению или предотвращению инициации трансляции данных кодирующих последовательностей мРНК рибосомами; именно этим путем достигается избирательная или преимущественная трансляция одних мРНК или отдельных ее цистронов и выключения трансляции других. Кроме того, разная скорость инициации на разных мРНК или их цистронах определяет соотношение продукции соответствующих белков.
В связи со сказанным, существует специальный, довольно сложный механизм инициации трансляции, обеспечивающий как точное узнавание начала кодирующей последовательности и ее дальнейшую фазировку, так и возможность положительных и отрицательных регуляторных влияний.
Инициирующие кодоны. Трансляция природных мРНК или ее отдельных цистронов (в случаях полицистронных мРНК) начинается, как правило, с триплета AUG или, гораздо реже, GUG или UUG. Эти триплеты, таким образом, могут служить как инициирующие кодоны матрицы. В некоторых, еще более редких, случаях триплеты AUU и AUA также, по-видимому, могут быть инициирующими кодонами. Таким образом, инициирующий кодон, - как правило, AUG - начальная точка отсчета триплетов вдоль матрицы по направлению к ее 3'-концу,
Инициаторная тРНК. Инициирующие кодоны узнаются специальной инициаторной тРНК. Антикодоном этой тРНК служит CAU, который способен спариваться с инициирующим кодоном, как правило, с AUG, но также и с GUG,UUG, AUU или AUA, когда они являются инициирующими.
Белковые факторы инициации. Существуют три специальных белка, необходимых для процесса инициации у прокариот; они получили название факторов инициации IF-1, IF-2 и IF-3.
IF-1 представляет собой небольшой белок основного характера с молекулярной массой около 9000 дальтон. IF-1 обнаруживается не у всех видов бактерии [7]. IF-2, наоборот, крупный белок кислой природы с важной для функции SH-группой. Это - главный фактор инициации. Он выделен в двух формах, несколько различающихся по молекулярной массе: одна (IF-2a) ~ около 100000, а другая (IF-2b) - около 90000 дальтон; обе формы, по-видимому, функционально эквивалентны в процессе инициации. IF-2 имеет сродство к ГТФ и образует с ним нестабильный комплекс. IF-2 с ГТФ взаимодействует с F-Met-tRNA и с рибосомой (с 30S субчастицей). ГТФ может быть заменен его нерасщепляемым аналогом [7]. IF-3 - слегка основного характера с молекулярной массой 21000-23000 дальтон;
для него описаны две функционально эквивалентные формы: одна — с длиной полипептидной цепи 181 аминокислотный остаток, и другая - укороченная на б остатков с N-конца [7].
Итак, во всех случаях инициации трансляции предшествует диссоциация нетранслирующих 70S рибосом на 30S и 50S субчастицы. Как уже говорилось, в физиологических условиях нетранслирующие рибосомы находятся в обратимом равновесии со своими субчастицами. IF-3 имеет довольно сильное сродство именно к диссоциированной 30S субчастице и, связываясь с ней, препятствует ее реассоциации с 50S субчастицей; таким образом, 30S субчастица уводится из равновесия и подготавливается для инициации. Два других фактора инициации, IF-1 и IF-2, также имеют большее или меньшее сродство к 30S субчастице и рдогут связываться с ней:
30SIF-3 + IF-1 + IF-2 <=> 30SIF-3IF-2IF-1
Далее, 30S субчастица с IF-3 или со всеми факторами инициации может ассоциировать с инициаторным районом мРНК. Этот район может находиться близко у 5'-конца или далеко от него, а в случае полицистронных мРНК таких районов может быть несколько вдоль всей цепи мРНК; важно лишь то, чтобы он был открыт для взаимодействия, имел подходящую конформацию и содержал прединициирующую полипуриновую последовательность Шайна—Дальгарно и инициирующий триплет AUG (или GUG, или UUG, и т. д.). Сама 30S субчастица, без факторов инициации, также способна узнавать инициирующий район мРНК и связываться с ним. Вместе с тем, IF-3, по-видимому, способствует ассоциации 30S субчастицы с мРНК, возможно, путем усиления комплементарного взаимодействия 3'-конца рибосомной РНК с последовательностью Шайна—Дальгарно или путем привнесения дополнительного взаимодействия между рибосомной частицей и матрицей.
На следующей стадии вступает в действие IF-2, либо уже ассоциированный с 30S субчастицей, либо находящейся в растворе. Он взаимодействует с ГТФ и с инициаторной F-Met-tRNAp, в результате чего имеет место кооперативное образование инициаторного 30S-KOMnneKca с освобождением IF-3:
30S mRNA • IF-3 + IF-2 + GTP + F-Met-tRNAp
30S • mRNA F-Met-tRNAp IF-2 • GTP + IF-3
Здесь, как и в случае факторов элонгации, ГТФ выступает в роли эффектора, придающего фактору IF-2 повышенное сродство к рибосоме (в данном случае к 30S субчастице) и к соответствующей тРНК (F-Met-tRNAp). По-видимому, сначала ГТФ взаимодействует с IF-2, а затем комплекс IF-2 • GTP связывает F-Met-tRNA; F-Met-tRNA узнает инициирующий кодон на 30S субчастице, a IF-2 • GTP обеспечивает дополнительное взаимодействие, комплексируясь одновременно с F-Met-tRNA и с рибосомной частицей. В процессе образования этого инициаторного 30S-комплекса ГТФ не расщепляется и, как уже отмечалось, может быть с полным успехом заменен на его нерасщепляемый аналог — гуанилилметилендифосфонат (GMPPCP) или гуанилилимидодифосфат (GMPPNP).
Однако не исключен и альтернативный путь образования инициаторного 308-комплекса. Дело в том, что рибосомная 30S субчастица без мРНК тоже имеет собственное сродство к инициаторной F-Met-tRNA, а также к IF-2. Это создает хорошую возможность взаимодействия F-Met-tRNA • IF-2 • GTP с 30S .субчастицей в отсутствие матрицы:
30S • IF-3 + IF-2 + GTP + F-Met-tRNAp
30S • F-Met-tRNAF IF-2 • GTP (+1F-3?)
Лишь после этого 30S субчастицы вместе с F-Met-tRNA специфически связываются с инициирующим районом и инициирующим кодоном мРНК;
антикодон F-Met-tRNA способствует специфичности и точности узнавания района с инициирующим триплетом на мРНК. Хотя в настоящее время большинство исследователей, работающих с бесклеточными системами, отдают предпочтение первому пути, не исключено, что в клетке сосуществуют оба пути, или даже второй может использоваться предпочтительнее.
Роль третьего, самого маленького, белка IF-1 не очень ясна в процессе образования инициаторного 30S-KOMnneKca. С одной стороны, есть указания на его вклад в увеличение скорости диссоциации нетранслирующих 70S рибосом на субчастицы. С другой стороны, он стабилизирует связывание двух других факторов инициации, IF-3 и IF-2, с 30S субчастицей, связываясь и сам в их присутствии (кооперативное связывание трех факторов инициации). IF-1 присутствует в конечном инициаторном 30S-KOMnneKCe с мРНК и F-Met-tRNA (в то время как IF-3, по-видимому, освобождается при связывании F-Met-tRNA) и, как полагают, стабилизирует его-
Инициаторный 30S-KOMnneKC теперь имеет сильное сродство к свободной 5 OS-субчастице, так что они ассоциируют, давая в результате 70S-комплекс. При этом F-Met-tRNA оказывается в Р-участке рибосомы. IF-2, ассоциированный с F-Met-tRNA и с 30S субчастицей, взаимодействует также с 50S субчастицей в комплексе. В результате взаимодействия наводится ГТФазная активность, и ГТФ гидролизуется на ГДФ и ортофосфат. Это, в свою очередь, приводит к резкому уменьшению сродства IF-2 к F-Met-tRNA и к рибосоме, так что IF-2 с ГДФ освобождаются в раствор. Имеются указания, что IF-1 сильно способствует освобождению IF-2 после распада ГТФ. 70S рибосома остается с мРНК и с инициаторной F-Met-tRNA в Р-участке, где инициирующий кодон связан с антикодоном инициаторной тРНК. Таким образом,
30SmRNA-F-Met-tRNAIF-2GTPIF-l+50S
<=>70SmRNAF-Met-tRNAIF-2GTPIF-l =>70SmRNAF-Met-tRNA+IF-2+GDP+Pi+IF-l
(существует альтернативное предположение, что IF-1 выходит из инициаторного комплекса после ассоциации с 50S субчастицей и до гидролиза ГТФ). Образовавшийся конечный инициаторный 70S-KOMiuieKC способен воспринять аминоацил-тРНК (с БР-Тц с ГТФ) на свой вакантный А-участок и начать элонгацию (см. схему 3).
Схема 3.
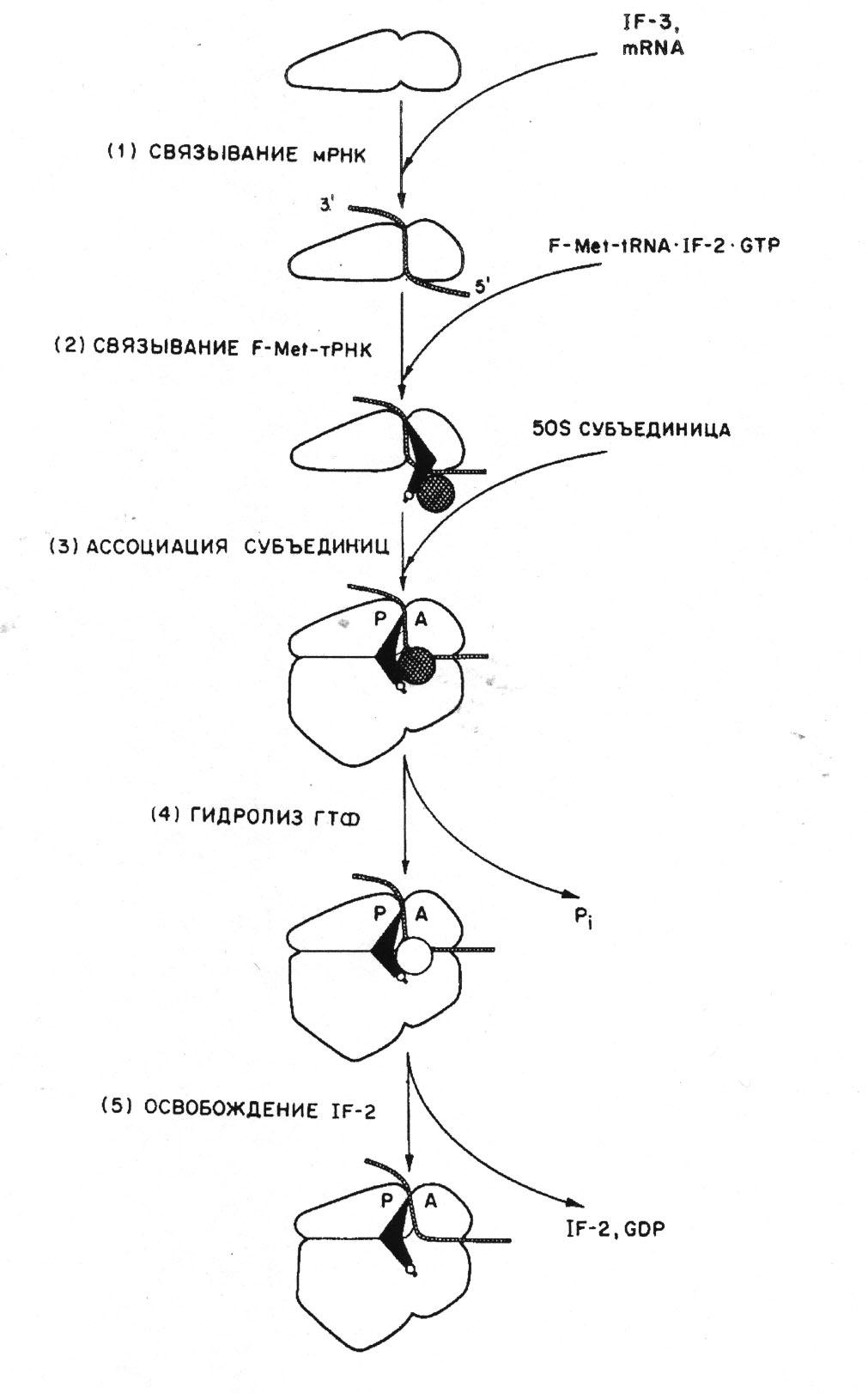
Схема последовательности событий в процессе инициации трансляции у прокариотических организмов 7.