

№1.
МИКРОБИОЛОГИЯ - наука о живых объектах, не видимых невооруженным глазом (с греческого - micros - малый; bios - жизнь; logos - наука).( бактерии,археи, микроскопическиегрибыиводоросли, часто этот список продляютпростейшимиивирусами)
Микробиология в настоящее время по праву может считаться одной из основных дисциплин биологии, поскольку без знаний особенностей микроорганизмов нельзя понять всего многообразия жизни на Земле, условий ее появления и эволюции. Микробиология имеет большое значение для многих наук, таких как, биофизика, генетика, молекулярная биология, экология и др.
История развития микробиологии: описательный, физиологический этапы.
Микроорганизмы были открыты и описаны в конце XVII века в Голландии Антонием ван Левенгуком. Сделанные им приборы — микроскопы давали по тем временам довольно сильное (более чем в 160 раз) увеличение.
Конец XVIII — начало XIX века в развитии микробиологии можно охарактеризовать как период описательный, собирательный. В это время были открыты и описаны различные микроорганизмы: дрожжи, вызывающие сбраживание пивного сусла (1837), паразитические грибы — возбудители болезни шелковичных червей (мускардины). В середине XIX века появились первые сообщения Поллендера и Брауэля в России (1848), Давена в Западной Европе о находках неподвижных нитевидных телец в крови животных, больных сибирской язвой. Однако только гениальные работы Пастера положили начало развитию современной научной микробиологии. Особенно велики заслуги Пастера в изучении сибирской язвы и бешенства. Его работы по созданию невосприимчивости к инфекционным заболеваниям были заложены основы вакцинопрофилактики, дающей блестящие результаты до настоящего времени.
ВТОРАЯ ПОЛОВИНАXIX ВЕКА
И. И. Мечников является создателем фагоцитарной теории иммунитета. Он показал, что одним из важнейших механизмов, помогающим человеку бороться с проникшими в его организм болезнетворными микробами, является клеточная защита. И. И. Мечников установил, что белые кровяные тельца — лейкоциты — захватывают и пожирают микробов, проникших в ткани человеческого организма. На месте проникновения микробов развивается воспалительная реакция, а гной — это погибшие лейкоциты. Клетки, пожирающие микробов, И. И. Мечников назвал фагоцитами (от греч. phagos — пожирающий, kytos — клетка). Роберт Кох — современник Пастера — внес огромный вклад в развитие медицинской микробиологии, открыл и изучил возбудителей таких тяжелых инфекционных заболеваний человека, как туберкулез и холера. Микробиологическая наука обязана Коху совершенствованием методов микробиологической, техники: он предложил способы окраски микроорганизмов, которые помогли изучить строение многих микробов, использовал при микроскопии освещение (осветитель Аббе), ввел микрофотографирование. Методы микробиологических исследований, разработанные Кохом, позволили получить чистую культуру возбудителей инфекционных болезней (микроорганизмы только одного вида). Это стало возможным при выращивании микроорганизмов на плотных питательных средах, предложенных Кохом. с 1891года Эрлих стал разрабатывать методы леченияинфекционных болезнейс помощью химических веществ. Он установил факт приобретениямикроорганизмамиустойчивости к химиотерапевтическим препаратам. Мировую славу Эрлиху принес разработанный им «препарат 606» (сальварсан), который оказался высокоэффективным при лечениисифилиса.
ПЕРВАЯ ПОЛОВИНАXX ВЕКА
Особые организмы, вызывавшие болезнь, — вирусымозаичной болезни табака — удалось увидеть впервые только в 1939 году вэлектронный микроскоп. Однако именно 1892 год считается годом открытия этих новых организмов — вирусов. Ивановский положил началовирусологии, выросшей в самостоятельную область науки.
C. Н. Виноградский (1856—1953) установил роль микроорганизмов в биологически важных процессах круговорота веществ в природе. Он разработал оригинальный метод накопительных культур, предложив селективные питательные среды, позволившие ему выделить и изучить аутотрофные микроорганизмы почвы: нитрофицирующие и азотфиксирующие.
Беспорядок в лаборатории Флеминга сослужил ему службу. В 1928 годуон обнаружил, что на агаре в одной из чашек Петри с бактериямиStaphylococcusвыросла колония плесневых грибов. Колонии бактерий вокруг плесневых грибов стали прозрачными из-за разрушения клеток. Флемингу удалось выделить активное вещество, разрушающее бактериальные клетки — пенициллин, работа была опубликована в1929 году.
|
Признак |
Прокариотная клетка |
Эукариотная клетка |
|
Организация генетического материала |
нуклеоид (ДНК не отделена от цитоплазмы мембраной), состоящий из одной хромосомы; митоз отсутствует |
ядро (ДНК отделена от цитоплазмы ядерной оболочкой), содержащее больше одной хромосомы; деление ядра путем митоза |
|
Локализация ДНК |
в нуклеоиде и плазмидах, не ограниченных элементарной мембраной |
в ядре и некоторых органеллах |
|
Цитоплазматические органеллы |
отсутствуют |
имеются |
|
Рибосомы в цитоплазме |
70S-типа |
80S-типа |
|
Цитоплазматические органеллы |
отсутствуют |
имеются |
|
Движение цитоплазмы |
отсутствует |
часто обнаруживается |
|
Клеточная стенка (там, где она имеется) |
в большинстве случаев содержит пептидогликан |
пептидогликан отсутствует |
|
Жгутики |
нить жгутика построена из белковых субъединиц, образующих спираль |
каждый жгутик содержит набор микротрубочек |
Схема пяти царств живого мира: прокариоты (царство Monera), одноклеточные эукариоты (царство Protista), многоклеточные эукариоты (царства Plantae, Fungi, Animalia) (по R. Whittaker, 1969)
Monera — прокариотные организмы, находящиеся на самом примитивном уровне
клеточной организации
Protista — микроскопические, одноклеточные, недифференцированные формы жизни
Plantae, Fungi, Animalia — растения (фототрофный тип питания ), грибы (питанием растворенными органическими веществами), животные (голозойное питание), т.е. многоклеточные эукариоты.
Современная классификация микроорганизмов. Домены: Bacteria, Archaea, Eukaria.
Микробы, или микроорганизмы (бактерии, грибы, простейшие, вирусы), систематизированы по их сходству, различиям и взаимоотношениям между собой. Этим занимается специальная наука — систематика микроорганизмов. Систематика включает три части: классификацию, таксономию и идентификацию. В основу таксономии микроорганизмов положены их морфологические, физиологические, биохимические и молекулярно-биологические свойства.
Различают следующие таксономические категории: царство, подцарство, отдел, класс, порядок, семейство, род, вид, подвид и др. В рамках той или иной таксономической категории выделяют таксоны — группы организмов, объединенные по определенным однородным свойствам.
Микроорганизмы представлены доклеточными формами (вирусы — царство Vira) и клеточными формами (бактерии, архебактерии, грибы и простейшие). Различают 3 домена (или «империи»): «Bacteria», «Archaea» и «Eukarya»:
домен «Bacteria» — прокариоты, представленные настоящими бактериями (эубактериями);
домен «Archaea» — прокариоты, представленные архебактериями;
домен «Eukarya» — эукариоты, клетки которых имеют ядро с ядерной оболочкой и ядрышком, а цитоплазма состоит из высокоорганизованных органелл — митохондрий, аппарата Гольджи и др. Домен «Eukarya» включает: царство Fungi (грибы); царство животных Animalia (включает простейшие – подцарствоProtozoa); царство растений Plante. Домены включают царства, типы, классы, порядки, семейства, роды, виды.
Штамм чистая культура микроорганизма, выделенная из определенного источника, и отличающаяся от других представителей вида.
Клон совокупность потомков, выращенных из единственной микробной клетки.
В настоящее время все большее признание получает систематика Берджи. Микроорганизмы, относящиеся к прокариотам, в определителе Берджи разделены на два отдела: I — циано-бактерии (сине-зеленые водоросли) и II — бактерии.
При изучении, идентификации и классификации микроорганизмов чаще всего изучают следующие (гено- и фенотипические) характеристики:
1. Морфологические - форма, величина, особенности взаиморасположения, структура.
2. Тинкториальные - отношение к различным красителям (характер окрашивания), прежде всего к окраске по Граму. По этому признаку все микроорганизмы делят на грамположительные и грамотрицательные.
3. Культуральные - характер роста микроорганизма на питательных средах.
4. Биохимические - способность ферментировать различные субстраты (углеводы, белки и аминокислоты и др.), образовывать в процессе жизнедеятельности различные биохимические продукты за счет активности различных ферментных систем и особенностей обмена веществ.
5. Антигенные - зависят преимущественно от химического состава и строения клеточной стенки, наличия жгутиков, капсулы, распознаются по способности макроорганизма (хозяина) вырабатывать антитела и другие формы иммунного ответа, выявляются в иммунологических реакциях.
6. Физиологические- способы углеводного (аутотрофы, гетеротрофы), азотного (аминоавтотрофы, аминогетеротрофы) и других видов питания, тип дыхания (аэробы, микроаэрофилы, факультативные анаэробы, строгие анаэробы).
7. Подвижность и типы движения.
8. Способность к спорообразованию, характер спор.
9. Чувствительность к бактериофагам, фаготипирование.
Размеры микроорганизмов.
Все живые объекты, размеры которых менее 70-80 мкм, невидимы невооруженным глазом и могут быть отнесены к микроорганизмам. Размеры микроорганизмов варьируют в широких пределах: величина самых крупных представителей приблизительно 100 мкм (некоторые диатомовые водоросли и высшие протисты), размеры одноклеточных зеленых водорослей и клеток дрожжей на порядок меньше приблизительно 10 мкм, еще меньше размеры, характерные для большинства бактерий. Типичная бактериальная клетка приблизительно 1 мкм в диаметре, в то время как большинство эукариотических клеток от 10 до 100 мкм в диаметре.
4. Форма микроорганизмов.
Прокариоты сферической формы (шаровидные клетки) называют кокками
Стрептококки (от греч. streptos цепочка) клетки округлой или вы тянутой формы, составляющие цепочку вследствие деления клеток в одной плоскости и сохранения связи между ними в месте деления. Сарцины (от лат. sarcina — связка, тюк) располагаются в виде пакетов из 8 и более кок ков, так как они образуются при делении клетки в трех взаимно перпендикулярных плоскостях. В этом случае колонии бактерий имеют правильную сферическую форму. Если деление происходит в нескольких плоскостях неравномерно, образуются клеточные скопления неправильной формы. Стафилококки (от греч. staphyle виноградная гроздь) представляют собой кокки, расположенные группами (гроздьями) в результате деления в раз ных плоскостях.
Прокариоты, имеющие форму цилиндра, палочковидные бактерии
Слегка изогнутые палочки называют вибрионами
Спириллы извитые формы бактерий, имеют вид штопорообразно извитых клеток
Спирохеты тонкие, длинные, извитые (спиралевидной формы) бактерии, отличающиеся от спирилл подвижностью, обусловленной «сгибательными» изменениями клеток.
Актиномицеты — ветвящиеся грамположительные бактерии
№2.
1. Ядерная зона и генетический аппарат прокариотной клетки.
Прокариоты – это организмы, в клетках которых отсутствует оформленное ядро. Функции ядра выполняет нуклеоид (то есть «подобный ядру»); в отличие от ядра, нуклеоид не имеет собственной оболочки.
Основу генома прокариот составляют кольцевые молекулы ДНК: прокариотические хромосомы и плазмиды. Множество молекул ДНК образует две взаимосвязанные подсистемы: хромосомную и плазмидную.
Хромосомная подсистема прокариотического генома
Основу хромосомной подсистемы прокариотического генома составляет прокариотическая (бактериальная) хромосома (генофор), входящая в состав нуклеоида – ядерноподобной структуры.
Бактериальная хромосома представляет собой кольцевую двуспиральную правозакрученную молекулу ДНК, которая свернута во вторичную спираль. Вторичная структура хромосомы поддерживается с помощью гистоноподобных (основных) белков и РНК. Точка прикрепления бактериальной хромосомы к мезосоме (складке плазмалеммы) является точкой начала репликации ДНК (эта точка носит название сайта OriC). Бактериальная хромосома удваивается перед делением клетки. Репликация ДНК идет в две стороны от сайта OriC и завершается в точке TerC. Молекулы ДНК, способные себя воспроизводить путем репликации, называются репликоны.
Утипичных прокариот в неделящейся клетке имеется одна бактериальная хромосома. Поэтому прокариоты в целом являются гаплоидами
2. Плазмиды.
Плазмиды и эписомы - это небольшие фрагменты ДНК, отличающейся от основной массы ДНК. Они часто реплицируются вместе с ДНК хозяина, но не нужны для выживания его клетки.
Сначала было принято различать эписомы и плазмиды: эписомы внедряются в ДНК хозяина, а плазмиды - нет. К эписомам относятся F-факторы и так называемые умеренные фаги. Сейчас обе группы называют одним общим термином "плазмиды". Плазмиды широко распространены в природе, и в последние годы их считают внутриклеточными паразитами или симбионтами, устроенными еще проще, чем вирусы. Плазмиды придают своим клеткам-хозяевам целый ряд особых свойств. Некоторые плазмиды являются "факторами резистенции" (R-плазмиды, или R-факторы: от англ. R = resistance - устойчивость), т.е. факторами, придающими устойчивость к антибиотикам
3. Клеточная стенка грамположительных бактерий.
состоит исключительно из толстого слоя пептидогликана, плотно прилегающего к клеточной мембране и пронизанного тейхоевыми илипотейхоевыми кислотами.
4. Клеточная стенка грамотрицательных бактерий.
слой пептидогликана существенно тоньше, между ним и плазматической мембраной находится периплазматическоепространство( локализован целый ряд ферментов, компонентов электрон-транспортных цепей и систем секреции белков), а снаружи клетка окружена ещё одной мембраной
5. Необычные клеточные стенки прокариот
Некоторые скользящие бактерии (миксобактерии, флексибактерии) способны в процессе перемещения по твердому субстрату периодически менять форму клеток, например, путем изгибания, что говорит об эластичности их клеточной стенки, и в первую очередь ее пептидогликанового слоя. Электронно-микроскопическое изучение, однако, обнаружило у них клеточную стенку, типичную для грамотрицательных эубактерий. Наиболее вероятное объяснение гибкости клеточной стенки этих бактерий — чрезвычайно низкая сшитость ее пептидогликанового компонента.
Клеточные стенки метанобразующих архебактерий содержат пептидогликан особого химического строения. У других представителей этой группы клеточная стенка состоит исключительно из кислого гетерополисахарида, а у некоторых экстремально галофильных, метанобразующих и ацидотермофильныхархебактерий — только из белка.
Архебактерии с клеточной стенкой белковой природы не окрашиваются по Граму, остальные типы архебактериальной клеточной стенки дают грамположительную реакцию.
Прокариоты без клеточной стенки.
При воздействии определенными химическими веществами оказалось возможным получать в лаборатории из разных видов эубактерий формы с частично (сферопласты) или полностью (протопласты) отсутствующей клеточной стенкой. Впервые это обнаружили при действии на бактериальные клетки лизоцимом, ферментом из группы гликозидаз, содержащимся в яичном белке, слезной жидкости и выделяемом некоторыми бактериями.
Уникальность химического состава клеточной стенки прокариот, ее отличие от таковой эукариот сделали возможным создание и применение лекарственных препаратов, специфически действующих только на прокариотную клеточную стенку. На этом основано действие пенициллина и некоторых других антибиотиков, подавляющих разные этапы синтеза пептидогликана. Пенициллин, например, ингибирует образование связей между пептидными хвостами на этапе "сшивания" полимера, происходящего в клеточной стенке в процессе роста прокариотной клетки
6. Функции клеточной стенки прокариот.
Клеточная стенка прокариот выполняет разнообразные функции: механически защищает клетку от воздействий окружающей среды, обеспечивает поддержание ее внешней формы, дает возможность клетке существовать в гипотонических растворах. В первую очередь в этом "заслуга" пептидогликана.
Структурная дифференцировка клеточной стенки у грамотрицательных эубактерий, приведшая к формированию дополнительного слоя в виде наружной мембраны, значительно расширила круг функций клеточной стенки.
Прежде всего, это связано с проблемами проницаемости и транспорта веществ в клетку. Наружная мембрана имеет специфические и неспецифические каналы (поры) для пассивного транспорта веществ и ионов, необходимых клетке, т. е. осуществляет функции молекулярного "сита". Наружная мембрана также препятствует проникновению в клетку токсических веществ, что находит отражение в большей устойчивости грамотрицательных эубактерий (сравнительно с грамположительными) к действию некоторых ядов, химических веществ, ферментов и антибиотиков.
Разнообразные функции выполняют макромолекулы, локализованные частично или полностью на внешней стороне клеточной стенки, контактирующей с окружающей средой; это специфические рецепторы для фагов и колицинов; антигены (липополисахарид грамотрицательных эубактерий, тейхоевые кислоты грамположительных); макромолекулы, обеспечивающие межклеточные взаимодействия при конъюгации, а также между патогенными бактериями и тканями высших организмов.
7. ЦПМ.
ЦПМ — белково-липидный комплекс, в котором белки составляют 50 – 75 %, липиды — от 15 до 45 %. Кроме того, в составе мембран обнаружено небольшое количество углеводов. Как правило, липиды и белки составляют 95 % и больше вещества мембран.
Главным липидным компонентом бактериальных мембран являются фосфолипиды — производные 3-фосфоглицерина. Хотя у прокариот найдено множество различных фосфолипидов, набор их в значительной степени родо- и даже видоспецифичен.
Широко представлены в бактериальных мембранах различные гликолипиды. Стерины отсутствуют у подавляющего большинства прокариот, за исключением представителей группы микоплазм и некоторых бактерий. Так, в ЦПМ Acholeplasma содержится 10 – 30 % холестерина, поглощаемого из внешней среды, от общего содержания мембранных липидов. Из других групп липидов в мембранах прокариот обнаружены каротиноиды, хиноны, углеводороды.
Структура мембран
Мембранные липиды всех эубактерий и части архебактерий образуют бислои, в которых гидрофильные "головы" молекул обращены наружу, а гидрофобные "хвосты" погружены в толщу мембраны.
Углеводородные цепи, прилегающие к гидрофильным "головам", довольно жестко фиксированы, а более удаленные части "хвостов" обладают достаточной гибкостью. У некоторых архебактерий (ряд метаногенов, термоацидофилы) мембранные липиды, в состав которых входит C 40-спирт, формируют монослойную мембрану, по толщине равную бислойной. Монослойные липидные мембраны обладают большей жесткостью сравнительно с бислойной. При "биологических" температурах мембранные липиды находятся в жидкостно-кристаллическом состоянии, характеризующемся частичной упорядоченностью структуры. При понижении температуры они переходят в квазикристаллическое состояние. Чем более не насыщены и разветвлены остатки жирных кислот или чем большее число циклических группировок они содержат, тем ниже температура перехода из жидкостно-кристаллического состояния в квазикристаллическое.
"Жидкая" структура мембран обеспечивает определенную свободу молекул белков, что является необходимым для осуществления процессов транспорта электронов и веществ через мембрану. Это же свойство обусловливает высокую эластичность мембран: они легко сливаются друг с другом, растягиваются и сжимаются.
Функции ЦПМ прокариот
ЦПМ прокариот выполняет разнообразные функции, в основном обеспечиваемые локализованными в ней соответствующими ферментными белками. Первоначально была постулирована барьерная функция клеточной мембраны, получившая позднее экспериментальное подтверждение. С помощью специальных переносчиков, называемых транслоказами, через мембрану осуществляется избирательный перенос различных органических и неорганических молекул и ионов. В ней локализованы ферменты, катализирующие конечные этапы синтеза мембранных липидов, компонентов клеточной стенки и некоторых других веществ.
Общепризнана роль ЦПМ прокариот в превращениях клеточной энергии. У бактерий, источником энергии для которых служат процессы дыхания или фотосинтеза, в ЦПМ определенным образом расположены переносчики цепи электронного транспорта, функционирование которых приводит к генерированию электрохимической энергии (DmH+), используемой затем в клетке по разным каналам, в том числе и для образования химической энергии (АТФ).
8. Внутрицитоплазматические мембраны прокариот. Включения и запасные вещества.
В клетках разных групп прокариот обнаружены мембраны, построенные по принципу элементарной, иные, нежели ЦПМ. Среди внутрицитоплазматических мембран выделяют несколько видов. Развитая система внутрицитоплазматических мембран характерна для большинства фотосинтезирующих эубактерий. Поскольку было показано, что в этих мембранах локализован фотосинтетический аппарат клетки, они получили общее название фотосинтетических мембран. У некоторых организмов (пурпурные бактерии) фотосинтетические мембраны сохранили тесную связь с ЦПМ, легко обнаруживаемую при электронно-микроскопическом изучении ультратонких срезов клетки. У цианобактерий эта связь менее очевидна. Внутрицитоплазматические мембраны фотосинтезирующих эубактерий могут иметь вид трубочек, пузырьков (везикул, хроматофоров) или уплощенных замкнутых дисков (тилакоидов), образованных двумя тесно сближенными мембранными пластинами (ламеллами).
У прокариот, принадлежащих к разным группам, описаны локальные впячивания ЦПМ, получившие название мезосом. Хорошо развитые и сложно организованные мезосомы характерны для грамположительных эубактерий. У грамотрицательных видов они встречаются значительно реже и относительно просто организованы. Мезосомы различаются размерами, формой и локализацией в клетке. Выделяют три основных типа мезосом: ламеллярные (пластинчатые), везикулярные (имеющие форму пузырьков) и тубулярные (трубчатые).
Существуют разные точки зрения относительно роли мезосом в клетке. Согласно одной из них мезосомы не являются обязательной структурой, а служат только для усиления определенных клеточных функций, увеличивая общую "рабочую" поверхность мембран. Получены данные о том, что с мезосомами связано усиление энергетического метаболизма клеток. Мезосомы играют роль в репликации хромосомы и ее последующем расхождении по дочерним клеткам, участвуют в процессе инициации и формирования поперечной перегородки при клеточном делении. Для некоторых грамположительных бактерий обнаружено участие мезосом в секреторных процессах.
Запасные вещества прокариот. Некоторые цитоплазматические включения имеют явно приспособительное значение. И наконец, многие из них являются запасными веществами, отложение которых клеткой происходит в условиях избытка питательных веществ в окружающей среде, а потребление наблюдается, когда организм попадает в условия голодания. Проблема консервирования энергии решена прокариотами путем синтеза восстановленных высокополимерных молекул, главным образомполисахаридов, реже липидов или полипептидов. Молекулы запасных веществ плотно упакованы в гранулах и часто окружены белковой оболочкой. В таком виде они находятся в осмотически неактивном состоянии, что очень важно для клетки.
Запасные вещества прокариот представлены полисахаридами, липидами, полипептидами, полифосфатами, отложениями серы. Общими свойствами липидов является их нерастворимость в воде и растворимость в органических растворителях. У прокариот липиды входят всостав клеточных мембран и клеточной стенки, служат запасными веществами, являются компонентами пигментных систем и цепей электронного транспорта. Ниже мы рассмотрим синтез жирных кислот и фосфолипидов, являющихсяуниверсальным компонентом клеточных мембран
9. Цитозоль и рибосомы.
Содержимое клетки, окруженное ЦПМ, называется цитоплазмой. Фракция цитоплазмы, имеющая гомогенную консистенцию и содержащая набор растворимых РНК, ферментных белков, продуктов и субстратов метаболических реакций, получила название цитозоля. Другая часть цитоплазмы представлена разнообразными структурными элементами: внутрицитоплазматическими мембранами (если они есть), генетическим аппаратом, рибосомами и включениями разной химической природы и функционального назначения.
Рибосомы — место синтеза белка — рибонуклеопротеиновые частицы размером 15 – 20 нм. Их количество в клетке зависит от интенсивности процессов белкового синтеза и колеблется от 5000 до 90 000. Общая масса рибосом может составлять примерно 1/4 клеточной массы, а количество рибосомальной РНК (рРНК) — 80 – 85 % всей бактериальной РНК. Они построены из двух неодинаковых субчастиц.
Синтез белка осуществляется агрегатами, состоящими из рибосом, молекул информационной и транспортных РНК и называемыми полирибосомами, или полисомами. Последние могут находиться в цитоплазме или же быть связанными с мембранными структурами.
10. Капсулы, слизистые слои, чехлы.
Многие микроорганизмы продуцируют на поверхности клетки слизистое вещество. В зависимости от толщины слизистого слоя принято различать микрокапсулу толщиной до 0,2 мкм (она видима лишь в электронном микроскопе). Связь микрокапсулы с клеточной стенкой настолько прочна, что ее иногда предлагают рассматривать как элемент клеточной стенки. Макрокапсула представлена слоем слизи толщиной более 0,2 мкм. Слизью называют вещество, окружающее клетку, имеющее аморфный, бесструктурный вид и легко отделяющееся от поверхности бактериальной клетки, а по толщине часто превосходящее ее диаметр.
В большинстве случаев капсула образована полисахаридами. Капсулы же других видов состоят из полипептидов, представленных полимерами, в которых содержится много D- и L-форм глутаминовой кислоты.
Для ряда бактерий показана также способность синтезировать капсулу, состоящую из волокон целлюлозы.
Капсулы и слизи выполняют следующие функции:
• защитную – предохраняют клетку от действия различного рода неблагоприятных факторов внешней среды (механических повреждений, высыхания и т. п.);
• создают дополнительный осмотический барьер;
• способны выступать в качестве фактора вирулентности у некоторых бактерий;
• служат барьером для бактериофагов, препятствуя их адсорбции на клетках бактерий;
• являются источником запасных питательных веществ;
• объединяют клетки в цепочки, колонии;
• обеспечивают прикрепление клеток к поверхности субстрата.
Капсульные полисахариды, образуемые бактериями, имеют большое практическое значение. Ксантан, внеклеточный полисахарид бактерий Xanthomonascampestris, используется в составе смазок, при добыче нефти, в пищевой промышленности для улучшения вкусовых свойств консервированных и замороженных продуктов, соусов, кремов, а также в косметической промышленности.
В отличие от капсул и слизистых слоев, чехлы имеют сложную тонкую структуру; в их составе выявляют несколько слоев разного строения. Чехлы обычно имеют и более сложный химический состав. Например, чехол бактерий Sphaerotilisnatans содержит 36 % углеводов, 11 –гексозамина, 27 – белков, 5,2 – липидов и 0,5 – фосфора. Чехлы ряда бактерий, метаболизм которых связан с окислением восстановленных соединений металлов, часто инкрустированы их окислами. Следует отметить, что между капсулами, чехлами и слизистыми слоями у прокариот обнаружено много переходных форм, что часто не позволяет точно отличить капсулу от слизистых клеточных выделений или капсулу от чехла.
№3.
1. Покоящиеся формы прокариот.
Их общей морфологической особенностью являются утолщенные клеточные стенки, а также частое образование дополнительных поверхностных структур на фоне одновременной дегидратации и компактизации метаболического и генетического аппаратов. В результате этого покоящиеся клетки характеризуются чрезвычайно низким (фоновым) уровнем обмена веществ и энергии, но приобретают повышенную устойчивость к действию разнообразных повреждающих факторов: высоких и низких температур, кислотности среды, радиации и пр. При попадании же в благоприятные условия покоящиеся клетки прорастают с образованием активно метаболизирующих вегетативных клеток. Покоящиеся формы прокариот можно условно разделить на несколько видов: эндоспоры, экзоспоры, цисты, акинеты
Эндоспоры – это особый тип покоящихся клеток эубактерий, формирующихся внутри цитоплазмы родительской клетки и обладающих специфическими структурами: внутренней и наружной мембранами, кортексом и многослойными белковыми покровами. К спорообразующим относится большое число эубактерий, относящихся к 15 родам, и имеющих клеточную стенку типичную для грамположительных бактерий
Экзоспоры фототрофных бактерий формируются в результате отпочковывания от одного из полюсов материнской клетки. Они не имеют каких-либо особенных поверхностных структур, однако клеточная стенка значительно плотнее и толще аналогичной структуры вегетативной клетки. Экзоспоры миксобактерий - миксоспоры – формируются в т.н. «плодовом теле» псевдоплазмодия, клетки переходят в покоящееся состояние, их клеточная стенка утолщается и окружается капсулой. Цисты распространенный тип покоящихся клеток, образуемые представителями разных групп эубактерий, виды Azotobacter и скользящие бактерии. При истощении пищевых ресурсов в цисту превращается вся палочковидная клетка, а не только какая-то ее часть, что сопровождается изменением ее морфологии. Происходящие изменения включают потерю жгутиков, накопление в цитоплазме гранул поли-бета- гидроксимаслянной кислоты и образованием дополнительных клеточных покровов. Возникающие толстостенные шарообразные покоящиеся клетки – цисты – устойчивы к высушиванию, механическим нагрузкам и облучению, но не к дейтсвию высоких температур. Акинеты - это особая форма сохранения у планктонных нитчатых цианобактерий, обладающая повышенной устойчивостью к ряду пониженным температурам, высушиванию, но не к повышенным температурам. В акинету превращается вся вегетативная клетка, которая начинает этот процесс с увеличения своих размеров и накопленияв цитоплазме гранул запасных веществ (гликогеновых, полифосфатных, цианофициновых), а также карбоксисом при одновременном уменьшении количества хлорофилла и фикобилиновых пигментов. Пептидогликановый слой КС утолщается, окружающий слизистый чехол уплотняется за счет полисахаридных фибрилл. Под действием силы тяжести акиеты опускаются на дно водоемов и перезимовывают, а весной прорастают, после чего молодые, снабженные газовыми везикулами вегетативные клетки всплывают к поверхности воды.
2. Процесс споруляции, на примере формирования эндоспор у бактерий.
Образование эндоспор - процесс, имеющий место только в мире прокариот. Бактериальные эндоспоры - это особый тип покоящихся клеток грамположительных эубактерий, формирующихся эндогенно, т.е. внутри цитоплазмы"материнской" клетки (спорангия), обладающих специфическими структурами (многослойными белковыми покровами, наружной и внутренней мембранами,кортексом) и устойчивостью к высоким температурам и дозам радиации, летальным в норме для вегетативных клеток ( рис. 22, Г). Эндоспорам свойственно также и особое физическое состояниепротопласта.
К спорообразующим относится большое число эубактерий приблизительно из 15 родов, характеризующихся морфологическим и физиологическим разнообразием.. Среди них имеются палочковидные, сферические, мицелиальные формы, спириллы и нитчатые организмы. Все они имеют строение клеточной стенки, характерное для таковой грамположительных эубактерий. Ни в одном случае не выявлена наружная липополисахаридная мембрана, несмотря на то, что многие роды и виды спорообразующих бактерий не окрашиваются по Граму. По типу питания среди них обнаруженыхемоорганогетеротрофы,факультативныехемолитоавтотрофыи паразитические формы.
Отношение к кислороду также разнообразно: часть спорообразующих форм представлена аэробамии факультативнымианаэробами, другая часть включаетоблигатных анаэробов- от аэротолерантных форм до высокочувствительных к О2.
Первым шагом к спорообразованию является изменение морфологии ядерного вещества вегетативной клетки, образующего тяж вдоль длинной оси спорулирующей клетки. Приблизительно одна треть тяжа затем отделяется и переходит в формирующуюся спору. У некоторых видов ядерный тяж образуется только на одном полюсе клетки, в его формировании участвует не весь генетический материал вегетативной клетки, и впоследствии ядерный тяж целиком переходит в формирующуюся спору. Формирование споры начинается с того, что у одного из полюсов клетки происходит уплотнение цитоплазмы, которая вместе с генетическим материалом, представляющим собой одну или несколько полностью реплицированных хромосом, обособляется от остального клеточного содержимого с помощью перегородки. Последняя формируется впячиванием внутрь клеткиЦПМ. Мембрана нарастает от периферии к центру, где срастается, что приводит к образованию споровой перегородки. Следующий этап формирования споры - "обрастание" отсеченного участка клеточной цитоплазмы с ядерным материалом мембраной вегетативной клетки, конечным результатом которого является образованиепроспоры- структуры, расположенной внутри материнской клетки и полностью отделенной от нее двумя элементарными мембранами: наружной и внутренней по отношению к проспоре.
Между наружным и внутренним мембранными слоями проспоры начинается формирование кортикального слоя (кортекса). Затем поверх наружной мембраны проспоры синтезируются споровые покровы, состоящие из нескольких слоев, число, толщина и строение которых различны у разных видов спорообразующих бактерий. В формировании слоев споровых покровов принимает участие как наружная мембрана споры, так ипротопластматеринской клетки.
Все слои, окружающие протопласт эндоспоры, находятся внутри материнской клетки. На их долю приходится примерно половина сухого вещества споры. После сформирования споры происходит разрушение (лизис) "материнской" клеточной стенки и спора выходит в среду.
Спорообразование сопровождается активным синтезом белка. Белки эндоспор в отличие от белков вегетативных клеток богаты цистеином и гидрофобными аминокислотами, с чем связывают устойчивость спор к действию неблагоприятных факторов. Содержание ДНК в споре несколько ниже, чем в исходной вегетативной клетке, поскольку в спору переходит лишь часть генетического материала материнской клетки. Генетический материал поступает в спору в виде полностью реплицированных молекул ДНК. Споры некоторых видов содержат по 2 или 3 копии хромосомы. Содержание РНК в спорах ниже, чем в вегетативных клетках, и РНК в значительной степени при спорообразовании синтезируется заново. Помимо Са2+ в эндоспорах обнаружено повышенное содержание других катионов (Mg2+, Mn2+, К+), с которыми связывают пребывание спор в состоянии покоя и их термоустойчивость.
Существенные отличия эндоспор от вегетативных клеток выявляются при изучении химического состава отдельных споровых структур. Экзоспориумсостоит из липидов и белков и, вероятно, выполняет функцию дополнительного барьера, защищающего спору от внешних воздействий, а также регулирующего проникновение в нее различных веществ. Однако никаких данных, подтверждающих эти предположения, пока нет. Споровые покровы в основном состоят из белков и в небольшом количестве из липидов и гликолипидов. Белки покровов обладают высокой устойчивостью к неблагоприятным условиям и обеспечивают спорам защиту от действия литических ферментов, других повреждающих факторов, а также предохраняют спору от преждевременного прорастания. Оказалось, что споры мутантов, лишенные покровов, прорастают сразу же после выхода из материнской клетки, даже если условия для последующего роста неблагоприятны.
Кортекспостроен в основном из молекул особого типапептидогликана. При прорастании споры из части кортекса, прилегающей к внутренней споровой мембране, формируется клеточная стенка вегетативной клетки.
Жгутики бактерийсостоят из трёх субструктур:
Филамент (фибрилла, пропеллер) — полая белковаянить толщиной 10—20 нм и длиной 3—15 мкм, состоящая изфлагеллина, субъединицы которого уложены поспирали. Полость внутри используется при синтезе жгутика — он происходит в направлении отплазматической мембраны.
Крюк — более толстое, чем филамент (20—45 нм), белковое (не флагеллиновое) образование.
Базальное тело (трансмембранный мотор)
Базальное тело представляет собой систему колец, находящихся в ЦПМ и клеточной стенкебактерий. Два внутренних кольца — M и S-кольца (сейчас чаще рассматриваются как единое MS-кольцо) — являются обязательными элементами, причём M-кольцо находится в ЦПМ, а S — в периплазмеграмотрицательных и пептидогликановомслое грамположительных бактерий.
Синтез жгутика
Процесс синтеза жгутика эубактерий (Caulobactersp.) запускается экспрессией гена сtrA. Продуктом этого гена является белок CtrA. Синтез CtrA происходит сразу после перехода клетки из G0-фазы в S-фазу. Обычно участок ДНК, содержащий ген сtrA, метилирован. Синтезу белка CtrA предшествует деметилированиеДНК, которая затемреплицируется. После этого происходит синтез CtrA и егофосфорилированиекиназами.ГенсtrA имеет двапромотора: Р1 и Р2. С первого промотора осуществляется ингибирование синтеза CtrA. Со второго промотора осуществляется стимулирование синтеза CtrA. Следует отметить, что белок CtrA найден не у всех эубактерий, и механизм синтеза жгутика не универсален.
Механизм движения клетки
Вращение мотора вызывает пассивное вращение филамента. Болеемассивнаяклетка начинает вращаться примерно со скоростью 1/3 от скорости жгутика и в обратном направлении, а также приобретает поступательное движение.
У ряда бактерий мотор и жгутик могут вращаться только в одном направлении, переориентация происходит при остановке за счёт броуновского движения. Бактерии-перитрихи собирают при движении все свои жгутики (каждый из которых вращается против часовой стрелки) в один пучок. Для совершения кувырка они либо меняют направление вращения, либо изменяют его скорость, из-за чего пучок распадается. При полярном расположении жгутиков один из них может служить и толкающим, и тянущим приспособлением в зависимости от направления вращения.
Скорости движения бактерий варьируют от 20 мкм/с у некоторых Bacillus до 200 мкм/с у Vibrio. иному механизму у поверхности клетки.
Пили, фимбрии или ворсинки — поверхностные структуры, присутствующие у многих бактериальныхклетоки представляющие собой прямыебелковыецилиндры длиной 1—1,5мкми диаметром 7—10нм. Различаются по строению и назначению, причём у одной бактерии могут присутствовать несколько их типов. Во многих случаях функции пилей не до конца установлены, но всегда они так или иначе участвуют в прикреплении бактериальной клетки к субстрату.
Скольжение, скользящее движение — движение отдельных бактериальных клетокили их колоний по твёрдой поверхности вдоль их длинной оси без участия бактериальныхжгутиков.
№4. Метаболизм
Метаболизм ( обмен веществ ) – совокупность протекающих в клетке процессов, обеспечивающих воспроизводство биомассы.
Энергетический метаболизм – поток реакций, сопровождающихся мобилизацией энергии и преобразованием ее в электрохимическую или химическую (АТФ) форму, которая затем может использоваться во всех энергозависимых процессах.
Конструктивный метаболизм ( биосинтезы ) - поток реакций, в результате которых за счет поступающих извне веществ строится вещество клеток; это процесс связанный с потреблением свободной энергии, запасенной в химической форме в молекулах АТФ или других богатых энергией соединений.








Ассимиляция нитрата и нитрита

Ассимиляция аммония
![]()


![]()
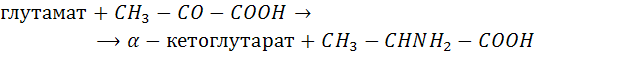
Азотфиксация – уникальный процесс, присущий только прокариотическим микроорганизмам, обладающих способностью к энзиматическому восстановлению атмосферного азота до аммония (с образованием водорода), который затем влючается в клеточное вещество.

Способы определения:
Ацетиленовый С2H2 + 2H+ → С2H4 CN− + 7H+ → NH3 + СH4 HN3 → NH3 + N2