

filo_ontogenez
.pdf29-го тижня це співвідношення поволі наростає, а розміри фолікулів поступово зменшуються. Індукуючий вплив на процес диференціювання тиреоїдного епітелію роблять фібробласти строми, що розвиваються з навколишньої мезенхіми.
Другим джерелом розвитку тиреоїдної паренхіми служать ультімобранхіальні тіла — парні формування, що формуються з
найбільш каудальних виростів дна ембріональної глотки. У риб вони є похідними VII глоткової кишені, у амфібій — VI, у рептилій і птахів — V—IV, у ссавців — IV (за винятком миші і щура, у яких є тільки 3 глоткових кишені). В ході ембріонального розвитку ультімобранхіальні тіла ссавців відділяються від матеріалу IV кишень, переміщаються каудально і закономірно інкорпоруються в долі медіального зачатка щитоподібної залози (у людини — на 7—8-му тижні).
Погляди на подальшу долю ультімобранхіальної тканини розходяться. Згідно однієї з найбільш відомих гіпотез, ця тканина підпадає під індукуючий вплив власне тиреоїдної паренхіми (що походить з медіального зачатка), хоча і зберігає свої потенції. Відповідно до другої гіпотези, вона руйнується, а згідно третьої — повністю диференціюється в типову тиреоїдну паренхіму, утворюючи інший, латеральний зарадок щитоподібної залози.
Найбільш обгрунтована перша гіпотеза. У щитоподібній залозі дорослих ссавців, включаючи людину, виявлені похідні ультімобранхіального тіла, що мають ряд характерних ознак. Друга гіпотеза, можливо, справедлива лише відносно якоїсь частини ультімобранхіального тіла. Можливість третьої гіпотези ставиться під сумнів результатами дослідів, які показали, що в експлантатах ультімобранхіальних тіл ембріонів, отриманих до їх злиття з медіальним зачатком, серед клітин, що диференціюються, типова тиреоїдна тканина відсутня.
Третім джерелом розвитку щитоподібної залози ссавців служить нервовий гребінь, який дає початок С-клітинам. У ембріо-
нальному розвитку малодиференційовані клітини нервового гребеня здатні здійснювати міграцію, що включає декілька етапів. На першому етапі відбувається виселення клітин з нервового гребеня у дорсо-вентральному напрямі у мезенхіму; на другому — вони проникають в епітелій IV глоткової кишені в області ультімобранхіального тіла, що формується. Разом з останнім вони далі мігрують каудально і, нарешті (також спільно з ультімобранхіальною тканиною), включаються в матеріал медіального зачатка щитоподібної залози, в якому розсіваються, перетворюючись на С-клітини. Невелика час-
241
тина С-клітин може диференціюватися поза зв'язком з ультімобранхіальним тілом за межами щитоподібної залози в інших похідних IV кишені — паращитоподібних залозах і тимусі.
Значення описаних процесів в ембріональному розвитку щитоподібної залози ссавців яскраво демонструється у випадках порушення нормального органогенезу, коли ультімобранхіальне тіло, що несе в своєму складі клітини нейрального походження, не зливається з медіальним зачатком залози. При цьому С-клітини, а також власне ультімобранхіальний епітелій диференціювалися у складі окремо лежачого ультімобранхіального тіла і не виявлялися у щитоподібній залозі. Такий варіант розвитку є нормальним для птахів,
рептилій, амфібій і риб.
Вікові особливості. Розміри щитоподібної залози у новонароджених значно більші, ніж у плодів. Протягом першого року життя відбувається зменшення маси щитоподібної залози, яка досягає 1,0 — 2,5 гр. До періоду статевого дозрівання розміри і маса поступово зростають (до 10—14 г). У період від 20 до 60 років маса органу залишається постійною і дорівнює в середньому 18 гр. Деяке зменшення маси і розмірів органу у зв'язку з віковою атрофією відбувається у старечому віці, проте функція щитоподібної залози у старості залишається непорушеною.
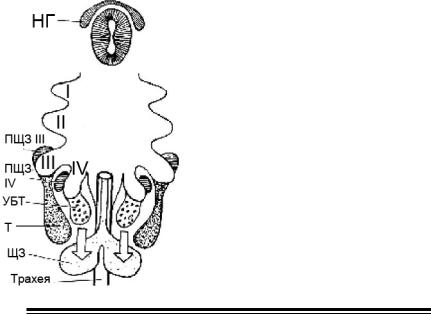
Мал. 19. Схема, що ілюструє ембріональний розвиток щитовидно- ї залози і інших залоз бранхіогенної групи (за Даніловим Р. Д.,
2001).
I, II, III, IV-відповідні зяброві кишені; НГнервовий гребінь; С-кл - С-клітини;
ПЩЖ III і ПЩЖ IV - паращитоподібні залози, що розвиваються з матеріалу відповідних зябрових кишень; УБТ -ультімобранхіальне тіло; Т - тимус; ЩЖ - щитовидна залоза.
242

Розвиток паращитоподібних залоз
Філогенез
Паращитоподібні залози виявляються у всіх класів хребетних, починаючи з риб. У хвостатих амфібій вони виявляються лише після метаморфоза. Передбачається, що поява паращитоподібних залоз у філогенезі пов'язана з виходом тварин на сушу, при якому у них виникла необхідність затримки кальцію в організмі. У водних тварин потреби в цьому не виникає, оскільки в морській воді концентрація кальцію вища, ніж в крові і тканинній рідині у риб, тоді як на суші вміст кальцію в їжі порівняно невеликий.
Улюдини і більшості тварин є дві пари паращитоподібних залоз, які по локалізації підрозділяють на верхні і нижні. Чотири паращитоподібні залози зустрічаються приблизно у 84% дорослих людей, більше число — у 3—13%, три — у 1—3%. Додаткові (ектопічні) паращитоподібні залози найчастіше пов'язані з тимусом. Іноді, навпаки, одна з пар паращитоподібних залоз відсутня. Таким чином, загальне число паращитоподібних залоз у людини може варіювати в межах від 2 до 12.
Ущурів, мишей і хом'яків в нормі є лише одна пара паращитоподібних залоз, яка розвивається з III зябрової кишені. Вони часто занурюються углиб долі щитоподібної залози в області латеральної поверхні верхнього полюса.
Онтогенез
Функціонально провідна тканина паращитоподібних залоз — залозистий епітелій — розвивається з матеріалу III і IV зябрових кишень, який, у свою чергу, походить з прехордальної пластинки, що має епідермальну детерміацію. За своїм початковим джерелом розвитку кожна пара паращитоподібних залоз позначається відповідно до найменування зябрової кишені, як паращитоподібні залози III і IV, відповідно. В області закладки залоз на 5-му тижні внутріутробного розвитку відбувається інтенсивне розмноження клітин, які утворюють компактні скупчення, що відділяються від зябрових кишень і зміщуються до місць своєї остаточної локалізації. Верхні паращитоподібні залози (IV) тісно зв’язані в своєму розвитку з ультімобранхіальним тілом. У міру того, як останнє в ході ембріонального розвитку включається в матеріал частки щитоподібної залози (у її задньолатеральній області), паращитоподібна залоза IV вступає
243

в тісний взаємозв’язок з щитоподібною залозою в цій ділянці, прикріпляючись до її капсули. Розташування верхніх паращитоподібних залоз відносно постійне; іноді вони занурюються углиб частки щитоподібної залози. Нижні паращитоподібні залози (III) спочатку лежать краніальніше паращитоподібних залоз IV. Їх остаточне більш каудальне і варіабельне розташування обумовлене активною міграцією на 7-му тижні внутріутробного розвитку у каудальному напрямі у тісному зв’язку з тимусом.
Паращитоподібні залози III зазвичай відділяються від тимуса і завершують свою міграцію в нижній частині шиї на рівні нижнього полюса щитоподібної залози. Область їх остаточного розташування — сполучна тканина латеральніше трахеї під нижніми полюсами доль щитоподібної залози. Розташування нижніх паращитоподібних залоз людини усередині залишків тимуса або навіть в середостінні пояснюється тим, що закладки залоз в ранньому ембріональному розвитку своєчасно не завершують свою міграцію у каудальному напрямі. Таким чином, у дорослої людини паращитоподібні залози розташовані у зворотньому порядку в порівнянні з місцями їх початкової закладки у ембріона.
На думку деяких дослідників, процеси, що сприймаються як міграція закладок залоз, насправді обумовлені не стільки їх активним переміщенням, скільки відмінностями швидкості росту сусідніх структур по відношенню до початкових ділянок формування паращитоподібних залоз.
У 5-місячного плоду вперше можна виявити темні і світлі паратироцити. Окремі жирові клітини в стромі з'являються перед народженням. Оксифільні клітини виявляються дуже рідко; фолікули, утворені паратироцитами, у плодів, як правило, не зустрічаються. Припускають, що паращитоподібні залози плоду на пізніх термінах
розвитку вже володіють ендокринною функцією.
Вікові особливості. Сумарна маса паращитоподібних залоз у новонародженого коливається від 6 до 9 міліграмів. Протягом першого року життя їх загальна маса збільшується в 3—4 рази, до 5 років вона подвоюється, а до 10 років потроюється. Після 20 років загальна маса паращитоподібних залоз досягає 120—140 міліграм і залишається постійною до глибокої старості. У всі вікові періоди маса паращитоподібних залоз у жінок дещо більша, ніж у
чоловіків.
Розвиток тимуса — див. розвиток центральних органів
імуногенезу у розділі 2.9
244

Розвиток надниркових залоз
Філогенез
Унижчих хребетних (круглоротих) інтерреналова тканина розташовується уздовж задніх кардинальних вен і ниркових артерій. Хромафінна тканина локалізується уздовж аорти, кардинальних вен
іінших судин.
Уриб інтерреналова і хромаффінна тканини знаходиться уздовж кардинальних вен або біля вен головної нирки, ці тканини формують одношарову або багатошарову муфту. До надниркових відносять особливі формування — тільця Станніуса.
Уземноводних надниркові залози виглядають у формі двох довгастих щілин неправильної форми, розташованих на вентральній поверхні нирок.
Уптахів острівці хромафінної тканини вкраплені серед інтерреналової тканини.
Уссавців кіркова речовина оточує мозкову.
Онтогенез
У ембріональному розвитку надниркова залоза формується з двох різнорідних зачатків, формуючих кіркову і мозкову речовину, які зливаються в єдиний орган. У ембріона людини на 4—5-му тижні розвитку відбувається проліферація целомічного епітелію в області кореня брижі біля краніального полюса первинної нирки, який вступає у взаємодію з мезенхімою і судинами, що розвивають-
ся. Ріст цих закладок, що продовжується, приводить до формування двох компактних утворень, покритих капсулою (інтерреналові тіла). В межах цих структур диференціюються дві зони: зовнішня,
яка складається з дрібних, щільно лежачих темних клітин і дає початок корі дефінітива дорослого організму, та внутрішня, яка пред-
ставлена крупними еозинофільними клітинами і утворює так звану фетальну кору. Остання у доношеного плоду складає близько 80%
всієї кіркової речовини, проте після народження піддається швидким дегенеративним змінам і зникає. Клітини фетальної кори мають ультраструктурні ознаки стероідпродукуючих елементів. Плацента і наднирники утворюють функціонально єдину систему. Фетальна кора регулюється АКТГ плоду; у атрофованих наднирниках аненцефальних плодів вона відсутня.
245

Кора дефінітива збільшується в об'ємі і диференціюється на три зони. Цей процес починається з появою клубочкової зони у 5-місячного плоду. На 6—7-му тижні розвитку поліпотентні клітини-попередники починають мігрувати з нейрального гребеня в епітеліальну закладку надниркових залоз, розмножуючись в ній,
і надалі накопичуючись в її центральній частині, утворюючи мозкову речовину. Диференціюючись в хромафінобласти, вони здатні
давати початок хромафінним клітинам, які складають основну масу мозкової речовини, а перетворюючись у симпатобласти — ган-
гліозним і підтримуючим клітинам. Компактні острівці хромафінних клітин («мозкові кулі»), що диференціюються, з'являються після 12-го тижня. Спочатку виявляються тільки клітини, що містять норадреналін, проте вже в процесі міграції у корі з'являються клітини, що синтезують адреналін. Передбачається, що програма розвитку клітин різних типів у мозковій речовині включається рядом чинників мікрооточення, які виробляються внутріутробно кірковою речовиною. Розвиток мозкової речовини у плоду відбувається відносно поволі і запізнюється у порівнянні з кірковою речовиною. Навіть до моменту народження мозкова речовина розвинена досить слабо.
В ході утробного розвитку наднирники швидко збільшуються у розмірах, переважно за рахунок росту фетальної кори, і у 3—4-місячних плодів вони більші за нирки. На пізніших термінах темп їх росту сповільнюється, а відносна маса знижується.
Частина інтерреналової тканини може розвиватися поза зв'язком з хромафінною тканиною у вигляді додаткових інтерреналових тілець у жировій тканині поблизу наднирників, у заочеревному просторі, в нирці (під капсулою), в органах статевої системи. Ці формування складаються виключно з клітин кіркової речовини, організованих за типом пучкової або сітчастої зон.
Мозкова речовина наднирників є частиною досить поширеної у людини хромафінної системи, яка дифузно розподілена по всьому організму. У значній кількості хромафінні клітини містяться в превертебральних і паравертебральних симпатичних гангліях, а також у так званих парагангліях — формуваннях, роз-
ташованих поблизу симпатичних вузлів. Найбільш крупний параганглій — черевний аортальний (орган Цукеркандля) — роз-
ташовується біля кореня нижньої брижової артерії. Він має до-
246

сить крупні розміри в ранньому ембріональному розвитку, проте з 8-го місяця внутріутробного життя по 1-й рік постнатально піддається інволюції. Характерно, що в парагангліях, на відміну від мозкової речовини наднирників, хромафінні клітини в ході розвитку не набувають здібності до синтезу адреналіну, а продовжують секретувати, як і в ембріональному періоді, тільки норадреналін. Цим побічно підтверджується вплив кіркової речовини
наднирників на диференціювання мозковогї.
Вікові особливості. Маса одного наднирника новонародженого значно перевищує масу наднирника дитини одного року життя. У перші три місяці після народження маса наднирника різко зменшується, головним чином за рахунок стоншення і перебудови кіркової речовини, а потім поступово відновлюється (до 5 років) і продовжує наростати. Остаточне формування кіркової речовини надниркових завершується у період другого дитинства. До 20 років маса кожного наднирника збільшується майже в 1,5 рази (в порівнянні з масою його у новонародженого) і досягає своїх максимальних розмірів. У подальші вікові періоди маса надниркових залоз практично не змінюється. Наднирники у жінок в середньому мають дещо більші розміри, ніж у чоловіків. Під час вагітності маса кожного наднирника збільшується приблизно на 2 гр. У пізні вікові періоди, після 70 років, спостерігається невелике зменшен-
ня маси і розмірів наднирників.
Розвиток ентодермальних залоз — острівкової частини підшлункової залози — див. розвиток травної системи.
Розвиток статевих залоз — див. розвиток статевої системи.
Розвиток дифузної ендокринної системи
Дифузну ендокринну систему (ДЕС) багато авторів називають APUD-системою (APUD — Amine Precursor Uptake and Decarboxylation). Але така назва, вказуюча на біохімічні особливості синтезу гормонів, не зовсім вірна, оскільки не всі клітини дифузної ендокринної системи синтезують гормони — похідні амінів. Тільки похідні шкірної ектодерми (див. нижче) є «дійсними» APUD-клітинами, оскільки синтезують гормони шляхом декарбоксилювання амінів. Ця плутанина у номенклатурі була викликана тим, що клітини дифузної ендокринної системи — похідні шкірної ектодерми, були відкриті першими, тому система і отримала назву APUD. Відкриття інших клітин ди-
247

фузної ендокринної системи відбулося порівняно недавно. Враховуючи морфологічні особливості будови системи (вона складається з окремих клітин, що синтезують різні гормони у різних органах і тканинах, причому ці клітини морфологічно не зв'язані між собою), логічно іменувати систему «дифузною ендокринною системою»
Згідно з сучасною класифікацією клітини дифузної ендокринної системи мають наступне походження:
1)похідні нейроектодерми — нейроендокринні клітини нейросекреторних ядер гіпоталамуса, епіфізу, мозкової речовини наднирників, пептидергичні нейрони центральної і периферичної нервової системи;
2)похідні шкірної ектодерми — клітини APUD-серії аденогіпофіза, клітини Меркеля в епідермісі;
3)похідні кишкової ентодерми – ентероцити, клітини гастроентеропанкреатичної (ГЕП) системи;
4)похідні мезодерми — секреторні кардіоміоцити, що розвиваються з міоепікардіальної пластинки;
5)похідні мезенхіми — огрядні клітини.
Аномалії розвитку ендокринних залоз
Аномалії розвитку залоз неврогенної групи — див. аномалії
розвитку нервової системи, розділ 2.10.
Аномалії розвитку залоз бранхіогенної групи (щитоподібна залоза, паращитоподібні залози, вилочкова залоза) — див. аномалії розвитку шиї.
Аномалії розвитку мезодермальних залоз (ендокринних частин статевих залоз) — див. аномалії розвитку чоловічої та жіночої статевих систем, розділ 2.6.
Аномалії ентодермальних залоз кишкової трубки (підшлункова залоза) — див. аномалії розвитку піжшлугкової залози, розділ 2.4.
Аномалії залоз адреналової системи — див. аномалії розвитку
нервової системи, розділ 2.10.
Аномалії розвитку щитоподібної залози
Наявність щитоязичної кісти, яка може бути розміщена у будь-якій ділянці на шляху міграції щитоподібної залози, але не дивлячись на це, завжди розташовується біля або ж
248

на серединній лінії шиї. Як видно з назви, кіста є залишком щитоязичної протоки. Хоча близько 50% цих кіст розташовано біля або під тілом під'язикової кістки, їх можна також знайти біля основи язика або щитоподібного хряща. Іноді щито-
язична кіста сполучається із зовнішнім середовищем за допомогою каналу — щитоязичної фістули. Така фістула зазвичай
виникає повторно, після прориву кісти, але може існувати і при
народженні.
Аберантну щитоподібну тканину можна знайти де завгодно уздовж шляху опускання щитоподібної залози. Переважно її знаходять в основі язика, відразу після сліпого отвору, і вона може ушкоджуватися тими ж патологічними процесами, що і сама залоза. У деяких випадках порушення процесу міграції закладки щитоподібної залози у каудальному напрямі приводить до розвитку всього органу або його частини краніальніше місця нормального розташування. Так, язична щитоподібна залоза розвивається в тих випадках, коли частина медіального зачатка дає початок тиреоїдній тканині в області початкової закладки органу. Зрідка в язичній залозі зосереджена вся функціонально активна тканина органу. У таких випадках її секреторна діяльність може бути недостатньою для забезпечення нормального метаболічного (еутіреоідного) стану.
Частою ембріональною аномалією щитоподібної залози є
розвиток краніально вираженої пірамідальної частки, що тягнеться до нижнього краю під'язикової кістки. У частині випадків зберігається своєрідна протока, що сполучає щитоподібну залозу із сліпим отвором; в ході нормального розвитку цей епітеліальний тяж піддається повному або переважному руйнуванню. Рідше уся щитоподібна залоза або її частина спускається каудальніше нормального розташування і опиняється в середостінні або входить в контакт з перикардом. Описаний випадок розташування тиреоїдної тканини у міжшлуночковій перегородці серця. У деяких осіб при посиленому розвитку додаткової тиреоїдної тканини основна щитоподібна залоза може бути відсутньою або залишатися недорозвиненою, у інших є як нормальна залоза, так і ектопічна тиреоїдна тканина. У патологічних умовах ектопічна тканина щитоподібної залози може розростатися, даючи незвично розташований зоб, а в деяких випадках — свищі, кісти і пухлини. Тому, знання її можливої топографії має велике клінічне значення.
249

2.9.РОЗВИТОК ОРГАНІВ КРОВОТВОРЕННЯ
ІІМУННОГО ЗАХИСТУ
Філогенез
Ухребетних тварин реакція імунного розпізнавання досягла особливої досконалості.
Круглороті — найдавніша група хребетних тварин. Характерно, що локальні, справжні осередки кровотворення з'являються у слизовій і підслизовій оболонках кишки, тому гемопоез невіддільний від лімфопоезу. Представникам цього класу, міногам і міксинам, властиві мононуклеарні клітини, здатні відторгати трансплантат. У них відбувається синтез антитіл до клітинних і білкових антигенів; вони дають специфічну відповідь після повторної імунізації бактеріофагами або еритроцитами, але кількість утворюваних антитіл дуже мала. Вироблення антитіл свідчить про появу специфічних В-клітин.
Уміксин відомий тільки один тип константної седиментації, близький до імуноглобуліну класу М. У міног ускладнюються імуноглобулінові молекули, у сироватці крові знайдено два види імуноглобулінів, подібних до таких класу IgМ і IgG ссавців. Імуноглобулінові молекули містяться в лімфоцитах периферичної крові. У міксин еволюція лімфоїдних структур пов'язана з становленням і розвитком видільної системи. Проте первинний алотрансплантат відторгується ще за хронічним типом. Так, у міксин він зберігає життєздатність близько 72 діб, а вторинний трансплантат — у 2,5 раза менше.
Уморських міног імунні реакції зумовлені лімфоїдними клітинами і рудиментарною селезінкою. У них вперше в еволюційному ряді виникає справжнє, не зв'язане з кровотворенням, морфологічне формування — тимус. Це дає підставу стверджувати, що у круглоротих виникає новий клітинний тип — Т-лімфоцит. Лімфоцити міног здатні до трансформації і поділу у відповідь на мітогени і специфічні антигени.
Риби характеризуються більш складною, у порівнянні з круглоротими, організацією лімфоїдної системи і досить сильною імунною відповіддю; мають повний імунний арсенал лімфоїдних органів. Вилочкова залоза, яка розміщується між другою і п'ятою зябровими пластинками, складається з часток і часточок. До лімфоїдних орга-
250