

6 курс / Эндокринология / Механизмы_нейроэндокринной_регуляции_Угрюмов_М_В_1999
.pdf
Рис. 23. Дни "рождения" нейронов, синтезирующих рилизинг-гормон гормона роста (РГГР), в аркуатном ядре у крыс,
определенные с помощью тимидиновой авторадиографии в К0мбинащ1и с имму-
ноцитохимией РГГР [Rodier et al., 1990]
По оси ординат - процент РГГР-имму- |
|
нореактивных меченных по [^Н]-тимидину |
|
нейронов в общей популяции РГГР-иммуно- |
|
реактивных нейронов, принятой за 100%. По |
|
оси абсцисс - дни введения [ЗН]-тимидина. |
|
1 - ростральная область аркуатного ядра; |
9 Ю^П 12 П 14 15 {"б 17 |
2 - средняя область аркуатного ядра; 3 - кау- |
Пренатальный период, дни |
дальная область аркуатного ядра |
|
ласть [De Gennaro et al., 1986; Ishikawa et al., 1986]. После этого распределение РГГР-нейронов становится таким же, как и у взрослых животных.
Характер выселения РГГР-нейронов в пренатальном периоде характеризуется видовыми особенностями. Так, у человека, в отличие от крысы, выселение нейронов начинается с латерального гипоталамуса (7-я неделя). Лишь спустя три месяца первые нейроны появляются в наиболее важном центре РГГР-системы - инфундибулярном ядре (аркуатном ядре), а затем и в других ядрах: паравентральном, перивентрикулярном, вентромедиальном и латеральном туберальном [Bresson et al., 1987b; Bugnon et al., 1987].
Дифференцировка РГГР-нейронов
Морфология. Малодифференцированные РГГР-нейроны плодов характеризуются небольшим размером, овальной формой и наличием одного или двух коротких отростков [Bloch et al., 1984; Daikoku et al., 1985]. Несмотря на то что по характеру распределения этих нейронов система выглядит вполне зрелой у крыс уже в раннем постнатальном периоде, а у человека к концу внутриутробного развития, сами нейроны еще в течение длительного времени выглядят малодифференцированными [Bloch et al., 1984; Ishikawa et al., 1986]. Другими словами, дифференцировка РГГР-нейронов происходит позднее, чем дифференцировка ряда других пептидергических нейронов, например ГРГ-нейронов.
Экспрессия гена и синтеза препроРГГР. Из сопоставления времени начала образования РГГР-нейронов, их выселения в ядра гипоталамуса и появления мРНК и самого пептида следует, что экспрессия гена и экспрессия синтеза препроРГГР в дифференцирующихся нейронах синхронизированы [Jansson et al., 1987; Burgunder, 1991]. Более того, оба процесса совпадают по времени с выселением нейронов в область окончательной закладки. С начала экспрессии специфических синтезов в РГГР-нейронах у крыс в пренатальном периоде и по крайней мере до двух-трехмесячного возраста в гипоталамусе увеличивается общее содержание мРНК РГГР [Argente et al., 1991] и самого пептида (рис. 24) [Jansson et al, 1987], что может свидетельствовать об интенсификации процессов транскрипции и трансляции в процессе дифференцировки нейронов и/или о постепенном увеличении числа
51


гпава II
НЕЙРОНАЛЬНЫЕ СИСТЕМЫ, ПРОДУЦИРУЮЩИЕ "НЕЙРОГИПОФИЗАРНЫЕ " ГОРМОНЫ
ВАЗОПРЕССИНЕРГИЧЕСКАЯ СИСТЕМА
Организация и функциональное значение вазопрессинергической системы
у взрослых млекопитающих
Молекула и ген. Существование биологически активного соединения, обладающего сосудосуживающим и антидиуретическим действием, было предсказано еще в конце девятнадцатого - начале двадцатого столетия по результатам анализа соответствующей физиологической активности экстрактов задней доли гипофиза. Гормон с указанными свойствами, названный впоследствии ВП, был первым выделенным и синтезированным биологически активным пептидом [Du Vigneaud, 1956]. В дальнейшем оказалось, что ВП является лишь небольшим фрагментом молекулы-предшест- венника - препроВП, в состав которой, помимо ВП, входят сигнальная последовательность, белок-носитель нейрофизин II и гликопротеин (рис. 25) [North 1987; Jard, 1990; Poulain, Jard, 1992].
Ген молекулы препроВП кодирует мРНК, образованную 578 нуклеотидами. Функциональные пептидные последовательности - ВП, нейрофизин II и гликопротеин - закодированы тремя различными экзонами (см. рис. 25) [Jard, 1990]. Независимо от класса позвоночных, ВП представляет собой циклический нонапептид, причем у разных видов могут варьировать аминокислотные остатки в положении 3, 4 и 8. Так, у млекопитающих синтезируется аргинин-ВП (Ьу5*^-ВП), у немлекопитающих тетрапод - мезотоцин и аргинин-вазотоцин, а у рыб - аргинин-вазотоцин и изотоцин [Acher, 1984].
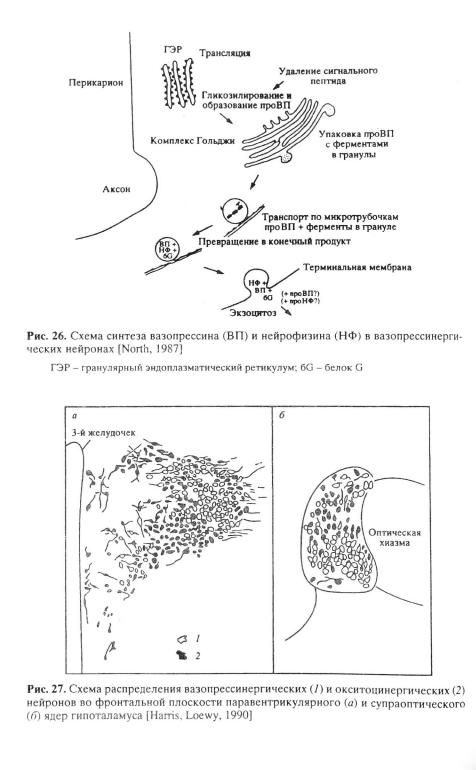
Крупноклеточные вазопрессинергические и окситоцинергические нейроны являются удобной моделью для изучения цитофизиологических аспектов синтеза нейрогормонов, их ферментативного преобразования и выделения. На этой модели впервые для нейросекреторных нейронов удалось локализовать на субклеточном уровне процессы синтеза, ферментативного превращения и выделения молекул препронейрогормонов и их производных. При этом оказалось, что трансляция мРНК с образованием белкапредшественника происходит на рибосомах гранулярного эндоплазматического ретикулума в телах и, возможно, в дендритах нейронов. После этого синтезированный белок поступает в цистерны гранулярного ретикулума
54
Рекомендовано к изучению сайтом МедУнивер - https://meduniver.com/
(рис. 26) [North, 1987], где он пре- |
|
Геи |
|
|
|
|||||
терпевает |
несколько последова- |
|
^ |
|
|
|
||||
тельных ферментативных преоб- |
|
|
|
|
|
|||||
разований: |
освобождение |
сиг- |
|
|
|
|
|
|||
нальной последовательности, гли- |
|
|
|
|
|
|||||
козилирование |
и |
образование |
Экзон 1 |
|
|
|
|
|||
восьми дисульфидных мостиков в |
|
|
|
|
|
|||||
молекуле препроВП. В таком ви- |
|
|
NH, |
|||||||
де полипептидные |
молекулы |
пе- |
Интрон А |
Х ч |
1 |
- |
19 |
|||
реносятся |
в комплекс Гольджи, |
|||||||||
X |
|
Сигнальный пептид |
||||||||
где происходят их окончательное |
|
|
|
|
Вазопрессин |
|||||
|
|
|
|
|
||||||
преобразование и упаковка в сек- |
|
|
|
|
|
|||||
реторные гранулы. В процессе по- |
|
|
|
|
|
|||||
следующей транспортировки гра- |
Экзон 2 |
|
|
|
Нейрофизин И |
|||||
нул в терминали аксонов в них |
|
|
|
|||||||
|
|
|
|
|
||||||
осуществляется |
ферментативное |
|
|
|
|
|
||||
расщепление препроВП с образо- |
|
|
|
|
|
|||||
ванием ВП, который выделяется |
Интрон В |
|
|
|
Гликопептид |
|||||
из аксонов в процессе экзоцитоза |
|
|
|
|||||||
гранул со всем остальным их со- |
|
|
|
|
145 |
|||||
держимым |
(см. рис. 26) [North, |
|
|
соон |
||||||
1987; Gainer et al., 1988]. |
|
|
|
|||||||
|
|
|
|
|
|
|||||
Архитектоника. |
Вазопрессин- |
Экзон 3 |
|
|
|
|
||||
ергическая |
система |
гипоталамуса |
|
|
|
|
||||
|
|
|
|
|
||||||
подразделяется на так называемые крупноклеточную и мелкоклеточную [Sofroniew, 1983]. Крупноклеточная система включает в себя вазопрессинергические нейроны относительно большого размера, которые располагаются в супраоптическом и паравентрикулярном ядрах (рис. 27), а также в так называемых добавочных ядрах гипоталамуса [Kozlowski et al., 1983]. Аксоны этих нейронов оканчиваются в
основном в задней доле гипофиза, где происходит выделение ВП в общую систему циркуляции [Falke, 1991].
Мелкоклеточная вазопрессинергическая система образована относительно мелкими неиронами, сосредоточенными в паравентрикулярном и супрахиазматическом ядрах. Большинство аксонов этих нейронов оканчивается в наружной зоне срединного возвышения, где ВП поступает в сосуды гипофизарной портальной системы циркуляции [Sofroniew, 1982]. Часть аксонов мелкоклеточных нейронов паравентрикулярного ядра проецируется на экстрагипоталамические отделы мозга - ствол мозга, включая белое пятно, солитарный парабрахиальный тракт и дорсальные ядра вагуса [Poulain, J^d, 1992], где они участвуют в синаптической иннервации нейро- нов-мишеней [Sofroniew, 1983].
55

Развитие вазопрессинергической системы
Происхождение вазопрессинергических нейронов
Несмотря на то что проведены десятки исследований развития вазопрессинергической системы у разных видов млекопитающих [Choy, Watkins, 1979; Whitnall et al., 1985; Laurent et al., 1989; Trembleau et a!., 1995; и др.], существует единственная работа, выполненная на мышах, в которой определены точные сроки образования вазопрессинергических нейронов гипоталамуса. В ней показано, что нейроны супраоптического и паравентрикулярного ядер образуются в одно и то же время - в первые дни второй половины внутриутробного развития [Okamura et al., 1983]. Наиболее вероятным источником этих нейронов являются нейроэпителиальные клетки ромбовидного расширения 3-го желудочка на уровне вентральной таламической бороздки [Altman, Bayer, 1978, 1986].
Первые нейроны, образующиеся из нейроэпителия, мигрируют в латеровентральном направлении в супраоптическое ядро [Fellmann et al., 1979; Whitnall et al., 1985; Bugnon et al., 1987; Yarnashita et al., 1988a, b]. Несмотря на то что начальная траектория миграции нейронов проходит через закладку паравентрикулярного ядра, в этот период развития нейроны там не задерживаются. Начиная от закладки паравентрикулярного ядра и до супраоптического ядра, нейроны мигрируют вдоль кровеносных сосудов, берущих начало от сосудов мягкой мозговой оболочки на уровне супраоптического ядра, врастающих в нервную ткань и восходящих по дуге в дорсомедиальном направлении.
Первые нейроны, экспрессирующие специфические маркеры - мРНК ВП и сам пептид, появляются в супраоптическом ядре одновременно с окончанием процесса образования вазопрессинергических нейронов из нейроэпителиальных клеток-предшественников, у мышей - на 15-16-й день развития [Yarnashita et al., 1988а, b; Hyodo et al., 1992]. Если это положение справедливо не только для незрелорождающихся, но и для всех остальных млекопитающих, то у человека процесс образования вазопрессинергических нейронов должен заканчиваться к 11-й неделе внутриутробного развития, когда в супраоптическом ядре впервые появляются ВП-имму- нопозитивные нейроны [Fellmann et al., 1979; Bugnon et al., 1987]. Использование общего маркера для вазопрессинергических и окситоцинергических нейронов - нейрофизина, позволило уточнить, что они начинают выселяться в супраоптическое ядро к концу 10-й недели внутриутробного развития, в добавочные ядра - на 13-й неделе и в паравентрикулярное ядро - на 14-й неделе [Mai et al., 1997]. Тем не менее, нельзя исключить, что дифференцирующиеся нейроны достигают супраоптического ядра еще раньше - до начала экспрессии специфических синтезов.
В процессе выселения из нейроэпителия 3-го желудочка вазопрессинергические нейроны сначала мигрируют в каудальную часть супраоптического ядра (рис. 28,1, II) [Buijs et al., 1980; Whitnall et al., 1985; Reppert, Uhl, 1987; Laurent et al., 1989; Buijs, 1992], a затем - до конца пренатального периода - распространяются в рострокаудальном направлении вдоль вентральной поверхности мозга от уровня оптических нервов и хиазмы до рет-
58
Рекомендовано к изучению сайтом МедУнивер - https://meduniver.com/