6 курс / Эндокринология / Механизмы_нейроэндокринной_регуляции_Угрюмов_М_В_1999
.pdf24 |
88 |
103 |
116 |
I |
I |
|
I |
S |
I |
^ |
Нетраяслкруеми |
i. |
5 |
I |
|
О |
область |
||
|
|
|
|
|
|
|
Экэон 1 |
Сигнальный |
COM (1-28) |
препро (46 ак) |
|
пептид |
|
|
|
Соматосгатин-14 |
|
|
|
Ala-Gly-(i^»-Ly»-Am-Ph&-Phe-TiT>-L)r»-Thr-Phe-Thr-Ser-Cyi |
Интрон = |
||
|
|||
Сомжтостатии-28 |
|
|
про (70 ак) |
Ser-Ala-Asn-Scr-Asn-Pro-Ala-Mct-AU-Pro-Arg-Oltt-Arg-Lye- |
Экэон 2 |
||
S |
|
S |
14С |]28 |
A1»-G1у-Суs-Lys~Asn—Phc-Phc-Trp-Lys-Thr-Phe-Thr-Scr-Cyi |
|||
Нетранслируемая
область
Рис. 16. Схематическое изображение препросоматостатина и фрагментов соматостатина (а), а также соответствующего им гена ((Т) [Kelly, 1990; Meister, Hokfelt, 1992]
COM - соматостатин, ак - аминокислота
Аксоны нейронов большинства ядер гипоталамуса, за исключением перивентрикулярного, не проецируются в срединное возвышение, а образуют сплетения в пределах ядер. При этом выделяющийся из них СС в качестве нейротрансмиттера и/или нейромодулятора влияет на нейроны-ми- шени [Liposits et al., 1988]. Обширное сплетение соматостатинергических волокон обнаружено также в сосудистом органе терминальной пластины, где они оканчиваются на сплетении капилляров общей системы циркуляции, а выделяющийся из них СС, по-видимому, оказывает влияние на висцеральные органы-мишени [Meister, Hokfelt, 1992].
У некоторых видов млекопитающих архитектоника соматостатинергической системы отличается от описанной выше. Например, у человека большая часть соматостатинергических нейронов сконцентрирована в паравентрикулярном ядре, в супрахиазматическом ядре и в латеральном гипоталамусе, а меньшая часть располагается в аркуатном, вентромедиальном и дорсомедиальном ядрах [Bouras et al., 1986, 1987].
Помимо гипоталамуса, СС-синтезирующие нейроны широко распространены в других отделах мозга: амигдале, гиппокампе, стриатуме, ядре солитарного тракта и спинальном ядре тройничного нерва, а также в периферической нервной системе - в верхних шейных ганглиях. Наряду с нервной системой, СС-продуцирующие железистые клетки широко представлены в ряде висцеральных органов, включая эндокринные железы (поджелудочная железа, щитовидная железа) и желудочно-кишечный тракт [Meister, Hokfelt, 1992; Konig, 1993].
39


Развитие соматостатинпродуцирующей системы
Происхождение соматостатинсинтезирующих нейронов
До настоящего времени среди млекопитающих только у крыс удалось определить время и место образования соматостатинергических нейронов, а также проследить пути их последующей миграции [Daikoku et al., 1983], Даже в этом случае был использован непрямой методический подход - иммуноцитохимия, позволяющая идентифицировать эти нейроны только после начала синтеза в них СС. В соответствии с упомянутыми выше эмбриогенетическими характеристиками были выделены три клеточные популяции. Нейроны наиболее ранней популяции образуются в начале второй половины внутриутробного развития (13-й день) в области нейроэпителия дна 3-го желудочка, граничащего каудально с преоптической бухтой. Со временем число этих нейронов возрастает, и вскоре (16-й день) они выселяются в аркуатное ядро [Daikoku et al., 1983].
Нейроны второй популяции образуются значительно позднее, чем нейроны первой популяции (18-й день), из нейроэпителия 3-го желудочка на уровне дорсальной области аркуатного ядра. В процессе дифференцировки эти нейроны мигрируют в дорсальный отдел аркуатного ядра и в вентральный отдел вентромедиального ядра.
Третья популяция образуется одновременно со второй, но в ростральном гипоталамусе из нейроэпителия боковых стенок 3-го желудочка [Daikoku et al., 1983]. Нейроны этой популяции выселяются в ростральную часть перивентрикулярного ядра [Shiosaka et al., 1992; Daikoku et al., 1983]. Последующее использование прямого методического подхода для определения времени образования соматостатинергических нейронов - комбинации тимидиновой авторадиографии с иммуноцитохимией - показало ограниченность только иммуноцитохимии для решения этого вопроса. Так, по данным, полученным с помощью двойного мечения, соматостатинергические нейроны перивентрикулярного ядра образуются гораздо раньше - с 13-го по 17-й день с максимумом на 14-16-й дни (рис. 18) [Hoffman et al., 1980], чем по результатам иммуноцитохимического исследования [Daikoku et al., 1983].
Выселение соматостатинергических нейронов не ограничивается упомянутыми вьппе областями гипоталамуса. Они также мигрируют в область мамиллоталамического тракта хронологически вслед за образованием соматостатинергических нейронов первой и третьей популяции, а также значительно позднее - к концу пренатального периода - в зону инсерта, латеральную преоптическую область, латеральный септум и в область диагонального пучка. Происхождение этих нейронов остается невыясненным [Shiosaka et al., 1992].
Число и размер соматостатинергических нейронов, по крайней мере выявляемых иммуноцитохимически, продолжают увеличиваться в гипоталамусе и в экстрагипоталамических отделах переднего мозга до конца внутриутробного развития и после рождения вплоть до периода полового созревания, что особенно ярко выражено в перивентрикулярном ядре - основном источнике аденогипофизотропного СС у взрослых животных
42
Рекомендовано к изучению сайтом МедУнивер - https://meduniver.com/
[Daikoku et al., 1983; Forloni et al., 1990]. Одним из редких исключений является латеральный септум, где количество соматостатинергических нейронов снижается после рождения [Shiosaka et
al., |
1992], что |
свидетельствует либо |
о |
дегенерации |
части нейронов, ли- |
бо о транзиторной экспрессии синтеза СС.
У человека, в отличие от других изученных млекопитающих, в развивающемся гипоталамусе были обнаружены не только мелкоклеточные соматостатинергические нейроны, но и крупноклеточные нейроны, синтезирующие наряду с СС препроВП и/или препроОТ [Bugnon et al., 1977 b, 1987]. Помимо размеров, нейроны этих двух популяций различаются по времени появления в гипоталамусе и локализации. Так, мелкоклеточные соматостатинергические нейроны появляются на 12-й неделе, будучи диффузно распределенными вокруг вентральной области 3-го желудочка на всем его протяжении от супрахиазматического ядра рострально до мамиллярных ядер каудально. Крупноклеточ-
14
312
10
ш о
3 4 5 6 7 Пренатальный период, дни
Рис. 18. Дни ''рождения" соматостатинергических нейронов (СС) у крыс, определенные с помощью тимидиновой авторадиографии в комбинации с иммуноцитохимией соматостатина [Hoffman et al,, 1980]
По оси ординат - процент сомато- статин-иммунореактивных меченных по [ЗН]-тимидину нейронов в общей популяции соматостатин-иммунореактивных нейронов, принятой за 100%. По оси абсцисс - дни введения [^Н]-тимидина
ные СС-содержащие нейроны выявляются с 16-й недели, располагаясь только в супраоптическом и пара-
вентрикулярном ядрах [Bugnon et al., 1977b, 1987].
Определенного внимания заслуживают сведения о том, что синтез СС в эмбриональном гипоталамусе не является прерогативой нейронов, а может осуществляться, по крайней мере, в культуре и глиальными клетками. При этом количество глиальных клеток, синтезирующих СС, зависит от нейрогуморальных факторов, входящих в состав плазмы культуральной среды [Davidson, Gillies, 1993].
Дифференцировка соматостатинергических нейронов
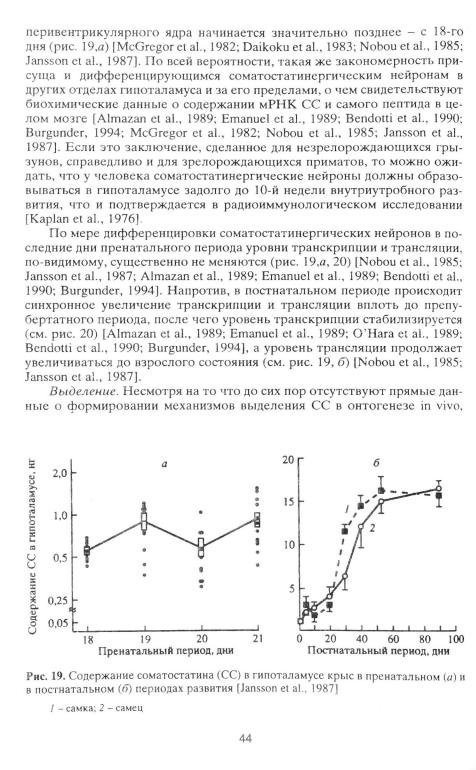
Экспрессия гена и синтеза препросоматостатина. Из сопоставления данных, полученных с помощью комбинации тимидиновой авторадиографии с иммуноцитохимией (см. рис. 18) [Hoffman et al., 1980], гибридизации in situ [Baram, Lemer, 1991; Baram, Schultz, 1991] и иммуноцитохимии [Daikoku et al., 1983], следует, что в нейронах наиболее важного с функциональной точки зрения перивентрикулярного ядра ген препроСС экспрессируется вскоре после их образования - у крыс начиная с 15-го дня пренатального периода. Однако синтез препроСС в нейронах
43