

2 курс / Нормальная физиология / Физиология_человека_Семенович_А_А_,_Переверзев_В_А_,_Зинчук_В_В
.pdf6.1. Функции нервной системы и ее структур
Общая характеристика функций нервной системы. Нерв! ная система регулирует жизнедеятельность клеток, тканей, органов и физиологических систем. Она участвует в регуляции гомеостаза и обеспечивает функциональное взаимодействие всех структур организма, их интеграцию, создает функцио! нальную целостность организма.
Благодаря нервной системе происходит адекватное взаимо! действие организма с окружающей средой. Это достигается на основе: 1) восприятия действия раздражителей из внешней и внутренней среды; 2) передачи информации о характере и силе раздражителей к нервным центрам; 3) анализа и синтеза по! ступающей информации. Синтез вновь полученной и накоплен! ной в процессе жизни информации обеспечивает принятие ре! шений и программы поведенческих реакций, необходимых для жизнедеятельности организма, а также сохранения социума.
Нервная система обеспечивает психические процессы, в том числе сознание, мышление, высшую познавательную и творческую деятельность.
Нервную систему подразделяют на центральную и перифе! рическую. Центральную нервную систему составляют голов! ной и спинной мозг. Мозг человека содержит около 25 млрд нервных клеток (нейронов). Скопление нервных клеток со! ставляет серое вещество ЦНС, а отростки этих клеток, объ! единяясь в проводящие пути, создают белое вещество. Кроме того, в ЦНС имеется нейроглия. Число глиальных клеток приблизительно в 10 раз превышает число нейронов, и эти клетки составляют большую часть массы ЦНС. Нейроны и глиальные клетки разделены межклеточными щелями (15– 20 нм). Эти взаимосвязанные межклеточные щели называют
интерстициальным пространством. Интерстициальное пространство занимает до 12–14% объема мозга.
Глиальные клетки служат опорным и защитным (входят в структуру гематоэнцефалического барьера) аппаратом для нейронов. Кроме того, нейроглия может влиять на возбуди! мость нейронов благодаря своей способности поглощать неко! торые ионы (особенно калия), выходящие в интерcтициальное пространство при высокой функциональной активности ней! ронов. Поглощая избыток ионов К+ из интерстициальной
131
жидкости, нейроглия снижает возбудимость нейронов, защи! щает их от возникновения судорожных разрядов.
К периферической нервной системе относят все нейроны, тела которых находятся вне ЦНС (в спинальных и вегетатив! ных ганглиях), и нервные волокна, выходящие за пределы го! ловного и спинного мозга. Такие волокна идут в составе череп! ных и спинномозговых нервов и нервных сплетений. К перифе! рической нервной системе относятся также окончания аффе! рентных волокон, образующие рецепторы, и окончания эфферентных нейронов, формирующие синапсы или подобные им образования (в частности варикозы окончаний постгангли! онарных вегетативных волокон). Важнейшими функциями пе! риферической нервной системы являются: 1) проведение воз! буждения (по афферентным волокнам в центростремительном направлении, по эфферентным – в центробежном); 2) генера! ция возбуждения в окончаниях афферентных нервных воло! кон, связанных с рецепторами; 3) передача возбуждения с эф! ферентных волокон через синаптические образования на эф! фекторы (мышечные волокна и тканевые структуры); 4) пере! работка информации нейронами, тела которых расположены в вегетативных ганглиях. Эта информация поступает к ним как от рецепторов, так и от центров головного и спинного мозга. Эти нейроны могут выполнять также функцию замыкания рефлекторной дуги так называемых периферических вегета!
тивных рефлексов.
Структура и функции нейронов. Нейрон (нервная клетка) является структурной и функциональной единицей ЦНС.
Анатомическая структура нейрона соответствует выполне! нию основных функций: получению, генерации и проведению нервных импульсов, переработке информации, передаваемой посредством нервных импульсов, объединению нейронов в нейронные цепи, обеспечивающие как простейшие рефлек! торные реакции, так и высшие интегративные функции мозга.
Нейроны имеют тело нервной клетки и отростки – аксон и дендриты (рис. 6.1).
Тело нейрона является морфологическим, трофическим и функциональным центром клетки. Здесь сосредоточено основ! ное количество нейроплазмы, ядро и клеточные органеллы, идет синтез веществ, необходимых для жизнедеятельности ак! сонов и дендритов. При повреждении тела отростки нейрона дегенерируют, разрушаются. Если же тело нейрона сохранено,
132

Рис. 6.1. Схема строения нейрона
а поврежден отросток, то происходит его восстановление, ре! генерация. Она идет медленно (несколько миллиметров в сут! ки). Вот почему при повреждении периферических нервов воз! можно восстановление иннервации мышц или органов.
Аксон – длинный (до 1,5 м) отросток нервной клетки. В месте выхода аксона из тела нейрона имеется утолщение – аксонный холмик – и далее идет непокрытый миелином учас! ток – начальный сегмент. Далее (у большинства нейронов) аксоны вплоть до своих конечных разветвлений покрыты мие! линовой оболочкой, прерываемой перехватами Ранвье – мик! роскопическими (около 5 мкм) безмиелиновыми участками.
Проницаемость и свойства мембраны, покрывающей тело и отростки нейрона, различаются. Наибольшей проницаемос! тью и вследствие этого наименьшей поляризацией (30– 50 мВ) обладает мембрана начального сегмента аксона. В бо! лее удаленных участках аксона величина трансмембранного потенциала составляет 70 мВ. Низкая поляризация начально! го сегмента аксона приводит к тому, что эта область нейрона обладает наибольшей возбудимостью.
При самых различных воздействиях на нейрон, вызываю! щих его деполяризацию, именно в области аксонного холмика, как правило, возникает потенциал действия, который прово! дится по аксону по направлению от тела нейрона к периферии.
В отличие от аксона дендритов на одном нейроне может быть несколько. Дендриты выполняют функцию передачи информации от периферии к телу нейрона. У большинства нейронов дендриты очень короткие и на них имеется много си!
133
наптических образований, через которые поступает поток ин!
формации от других нейронов.
Классификация нейронов. Проводится по морфологи! ческим и функциональным признакам.
Классификация и характеристика нейронов по морфологи! ческим признакам дана в учебнике анатомии (различают муль! типолярные, биполярные и псевдоуниполярные нейроны).
По морфофункциональному признаку выделяют: 1) аффе! рентные нейроны (их называют также чувствительными, цент! ростремительными); 2) вставочные, или ассоциативные, цент! ральные; 3) эфферентные, или двигательные, центробежные.
Афферентные нейроны проводят импульсацию от рецеп! торов к нервным центрам головного и спинного мозга. Поэто! му их часто называют также чувствительными или центростре! мительными. Тела этих нейронов находятся в спинальных и че! репно!мозговых ганглиях. Это псевдоуниполярные нейроны, аксон и дендрит которых отходят от тела нейрона вместе и за! тем разделяются. Дендрит идет на периферию к органам и тка! ням, а аксон входит в спинной или головной мозг.
Вставочные, или ассоциативные, нейроны выполняют функции переработки поступающей информации и, в частнос! ти, обеспечивают замыкание рефлекторных дуг. Тела этих нейронов располагаются в сером веществе головного и спин! ного мозга, а отростки не выходят за пределы ЦНС.
Эфферентные нейроны выполняют функцию передачи нервных импульсов от головного и спинного мозга к органам! эффекторам.
Имеются подразделения каждой из названных групп нейро! нов на подгруппы как по морфофункциональным, так и по функ! циональным признакам. Эти нейроны будут названы при рас!
смотрении конкретных выполняемых ими регуляций.
Особенности центральных синапсов. Важнейшие свой! ства и классификация синапсов были рассмотрены выше. Оста! новимся на особенностях центральных синапсов.
Центральные синапсы обеспечивают передачу информа! ции между нервными клетками, расположенными в головном и спинном мозге.
В зависимости от того, между какими отделами нейрона имеется синаптический контакт, синапсы подразделяют на ак! со!соматические (между аксоном одного и телом другого нейро! на), аксо!аксональные (между двумя аксонами), аксодендри!
134
тические (между аксоном и дендритом), дендро!дендритиче! ские (между двумя дендритами).
По эффекту влияния на активность нейронов синапсы под! разделяются на возбуждающие и тормозные. Активация тор! мозных синапсов приводит к снижению возбудимости или пре! кращению возбуждения нервных клеток.
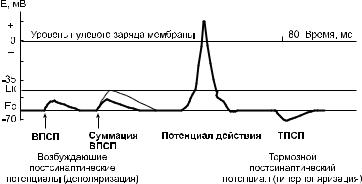
Структура и функционирование центральных синапсов имеют принципиальное сходство с рассмотренными выше нервно!мышечными синапсами, однако есть и некоторые раз! личия. На одном нейроне могут быть десятки тысяч синапсов. Одного потенциала действия, пришедшего по нервному волок! ну, недостаточно для возникновения возбуждения на эфферент! ном нейроне. Деполяризацию, возникающую на постсинапти! ческой мембране при активации центрального синапса, назы! вают возбуждающим постсинаптическим потенциалом – ВПСП (рис. 6.2).
Особенностью центральных синапсов является также раз! нообразие медиаторов (ацетилхолин, норадреналин, дофамин, серотонин, глютамин, аспарагин, глицин, ГАМК и др.), ис! пользуемых разными нейронами. Однако во всех синапсах од! ного нейрона используется один и тот же медиатор (принцип Дейла). Если им является ацетилхолин, то нейрон называют холинергическим, дофамин – дофаминергическим, норадре! налин – адренергическим и т.д.
Рис. 6.2. Возбуждающий и тормозной потенциалы, возникающие на постсинаптической мембране нейрона при активации соответствующих синапсов
135
В настоящее время выяснилась относительность этого принципа в связи с тем, что наряду с основным медиатором в синаптическую щель при активации синапса могут выбрасы! ваться сопутствующие нейромедиаторы, вещества пептидной природы, простагландины, эндорфины, энкефалины, АТФ. Эти вещества оказывают модулирующее действие на передачу возбуждения и могут проявлять долговременное действие на синаптические структуры, влияя на синтез и количество встра! иваемых в мембраны рецепторных белков, а также на энерге! тический обмен в нейроне.
Нервные импульсы, приходящие к нейрону, могут подав! лять его активность (вызывать торможение). Все виды тормо! жения подразделяют на 2 группы: первичное торможение и вторичное. Первичное торможение осуществляется через специализированные (тормозные) синаптические структуры. Его подразделяют на постсинаптическое и пресинаптическое.
Постсинаптическое торможение развивается в синапсах, ис! пользующих тормозные медиаторы (глицин, ГАМК и др.) и имеющих на постсинаптической мембране особые каналы (для ионов К+ и Сl–). Под действием тормозного медиаторы эти каналы открываются. Выход из клетки положительных заря! дов (с током ионов К+) и вход отрицательных (с током ионов Сl–) приводит к гиперполяризации постсинаптической мем! браны. Такую гиперполяризацию называют тормозным постси! наптическим потенциалом (ТПС П). Она и обусловливает сни! жение возбудимости нейрона. По признаку расположения вза! имодействующих нейронов постсинаптическое торможение подразделяют на прямое, возвратное, латеральное. При латеральном торможении возбужденный нейрон через боко! вые веточки своего аксона оказывает тормозное действие на рядом лежащие нейрона. Возвратное торможение характерно для мотонейронов. Веточка аксона такого нейрона активирует клетку Реньшоу, а она формирует тормозной синапс на теле мотонейрона (рис. 6.3). Примером прямого торможения мо! жет быть подавление активности конечного нейрона в тормоз! ных иерархических нервных цепях (рис. 6.3).
Пресинаптическое торможение реализуется путем ак! тивации аксо!аксональных синапсов. Тормозное влияние в этом случае происходит за счет деполяризации постсинапти! ческой мембраны. Например, так может блокироваться пере! дача на мотонейроны возбуждающих импульсов, проводимых по афферентным волокнам от рецепторов.
136
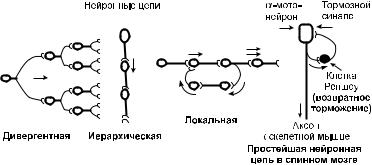
Рис. 6.3. Некоторые виды нейронных цепей
Для вторичного торможения не требуется наличия специа! лизированных структур. Один из его видов, называемый тор1 можение вслед за возбуждением, развиваться как результат предыдущего возбуждения. Так, у спинальных мотонейронов вслед за пиком ПД наступает длительный период гиперполя! ризации и в это время возбудимость их снижается. У нервных клеток имеется также универсальный тормозной механизм, называемый пессимальным торможением. Он срабатывает при возникновении в клетке чрезмерно частых волн возбужде! ния. При этом из!за длительной деполяризации происходит инактивация натриевых каналов и развитие новых волн воз! буждения блокируется. Такой механизм предохраняет клетку от чрезмерной активности, которая могла бы вызвать деструк! тивные изменения.
Процессы торможения – необходимое условие проявления координационной функции ЦНС.
Интегративная функция нейрона. Нейрон выполняет роль элемен! тарной функциональной единицы нервной системы благодаря воспри! ятию и переработке информации. Переработку информации и принятие решения называют интегративной функцией нейрона. Решение, прини! маемое нейроном, может выражаться в генерации и передаче определен! ной частоты возбуждений или в отсутствии возбуждения на нем. Возбуж! дение проявляется потенциалом действия.
Потенциал действия, как правило, возникает в области аксонного холмика, являющегося наиболее возбудимой частью нейрона. На аксон! ном холмике и начальном сегменте отсутствуют синапсы, но сосредото! чено большое число потенциалзависимых Na+!каналов. Основная масса синапсов сосредоточена на дендритах и теле нейрона. На дендритах их
137
число может достигать сотен тысяч, а на теле нейрона – 10 000. До 40% мембраны тела нейрона занято синаптическими структурами.
При активации возбуждающего синапса на постсинаптической мем! бране возникает ВПСП. Он распространяется с затуханием (на расстоя! нии 1–2 мм уменьшается на 37%) по телу нейрона и мембране началь! ного сегмента. Но величина одиночного ВПСП недостаточна для деполя! ризации этой мембраны до критического уровня и возникновения потен! циала действия. Он может образоваться лишь при большой частоте активации одного и того же синапса или одновременной активации мно! гих соседних возбуждающих синапсов. В этих условиях благодаря сумма! ции ВПСП область аксонного холмика деполяризуется до критического уровня и возникнет потенциал действия, передающийся далее от тела нейрона к окончаниям аксона.
Если одновременно с возбуждающими синапсами активируются тор! мозные, то они вызывают возникновение ТПСП. Эти гиперполяризую! щие потенциалы распространяются по мембране нейрона и по принципу алгебраической суммации снижают или полностью устраняют деполяри! зующий эффект ВПСП.
Таким образом, возникновение возбуждения на нейроне зависит от результата суммации на нем возбуждающих и тормозных постсинапти! ческих потенциалов. При этом основную роль играют постсинаптические потенциалы, возникающие на синапсах сомы. Меньшее значение имеют синапсы на дендритах, так как они удалены на значительное расстояние от места первичного возникновения ПД и мало влияют на поляризацию мембраны аксонного холмика.
Если к нейрону приходят активирующие импульсации, то на возник! новение возбуждения в этом нейроне необходимо время не менее 2 мс. Синаптическая же задержка составляет около 0,5 мс. Время, необходи! мое для возникновения возбуждения на эфферентном нейроне, может сильно удлиняться при увеличении активности тормозных синапсов на этом нейроне.
Нейронные цепи. Функциональные возможности нервной системы во многом обеспечиваются наличием нейронных цепей. К наиболее распространенным нейронным цепям от! носят: локальные, иерархические и дивергентные нейронные цепи с одним входом (рис. 6.3).
Локальные нейронные цепи могут выполнять функцию ловушек, в которых возбуждение способно длительное время бежать по кругу, образованному несколькими нейронами.
Возможность длительного существования однократно возникшей за счет движения по кольцевой структуре волны возбуждения впервые экспе! риментально показал проф. И.А. Ветохин в опытах на нервном кольце меду!
138
зы. Круговое движение возбуждения по нейронным цепям выполняет функ! цию трансформации ритма возбуждений, обеспечивает возможность дли! тельного возбуждения нервных центров после устранения раздражения, участвует в механизмах запоминания поступающей информации.
Локальные цепи могут выполнять также тормозную функ! цию. Это ярко видно на примере простейшей нейронной цепи в спинном мозге, образуемой α!мотонейроном и клеткой Рен! шоу (рис. 6.3).
Иерархические цепи обеспечивают связь и быструю пере! дачу управляющих сигналов между высшими и низшими отдела! ми нервной системы. Например, кора мозга управляет сокра! щением скелетных мышц за счет передачи команд по иерархи! ческим нервным цепям, обеспечивающим надежную и быструю передачу возбуждений к мотонейронам спинного мозга и череп! ным нервам. В такую цепь кроме нейрона моторной зоны коры могут входить нейроны ствола головного мозга и спинного мозга.
Дивергентные цепи с одним входом выполняют роль мно! жительной системы, передающей возбуждение от одного ней! рона сразу на многие нейроны. Это достигается за счет сильно! го ветвления (образования до 20 000 коллатералей) аксона. Такие нейроны часто встречаются в ядрах ретикулярной фор! мации ствола мозга. Они обеспечивают быстрое повышение возбудимости многочисленных отделов мозга и мобилизацию его функциональных резервов.
6.2. Свойства и принципы функционирования нервных центров
Свойства нервных центров. К ним относят:
1)замедленное распространение возбуждения, обу! словленное наличием синапсов и многих нейронов на путях пе! редачи возбуждения;
2)одностороннее распространение возбуждения, обусловленное тем, что основную роль в распространении воз! буждения играют химические синапсы, в которых возбуждение передается только от пресинаптических структур на постсинап! тические структуры;
3)иррадиация возбуждения – распространение возбуж! дения от активированного нервного центра на соседние и функ! ционально связанные нейронные структуры. Это свойство
139
обусловлено наличием многочисленных ветвлений аксонов, заканчивающихся синаптическими контактами. В результате при активации группы нейронов от них идут возбуждающие вли! яния на синаптически связанные с ними другие нейронные груп! пы. При прекращении притока возбуждающей импульсации активность нервного центра уменьшается и может наступать концентрация возбуждения – процесс, обратный иррадиации;
4)суммация возбуждения, проявляющаяся увеличением интенсивности рефлекторной реакции при возрастании дли! тельности раздражения, его силы или площади раздражаемого рецепторного поля. Различают временную и пространственную суммацию. При временной суммации количество активируемых синаптических образований на эфферентном нейроне не изме! няется, но увеличивается частота импульсаций, приходящих к каждому синапсу. При пространственной суммации из!за воз! действия на новые рецепторы увеличивается количество одно! временно активируемых синаптических входов и эфферентный нейрон быстрее и эффективнее возбуждается. В обоих случаях возрастает величина ответной реакции эффекторных структур. Например, увеличивается интенсивность сокращения мышцы, секреции слезной или слюнной железы.
5)трансформация ритма возбуждений, проявляющая! ся изменением частоты импульсаций, генерируемых эфферент! ными нейронами, по отношению к частоте импульсаций, пере! даваемых афферентными нейронами. Такое изменение стано! вится возможным благодаря суммации ВПСП, функциониро! ванию локальных нейронных цепей и т.д.;
6)явление последействия, заключающееся в способнос! ти нервных центров длительно поддерживать возбуждение после устранения раздражителя, вызвавшего его. Одним из механизмов этого служит возможность длительной циркуля! ции возбуждения в замкнутых нейронных цепях;
7)тонус нервных центров, проявляющийся длительным непрерывным возбуждением нервных центров. Такое состоя! ние поддерживается за счет непрерывного притока афферент! ных импульсаций, влияния биологически активных веществ, доставляемых гуморальным путем, циркуляцией возбуждения
взамкнутых нейронных цепях, спонтанной активностью от! дельных нейронов;
8)пластичность нервных центров – способность к пе! рестройке функциональных свойств и изменению выполняе! мых функций. Например, экспериментально показана способ!
140