

2 курс / Нормальная физиология / Физиология_дыхательной_системы_
.pdf«центром выдоха». Экспираторные нейроны разряжаются при выдохе и во время дыхательной паузы.
Таким образом, экспираторная область включает в себя ВЛГ нейронов, преимущественно каудальную часть. Она содержит:
1.Постинспираторные нейроны – интернейроны, активные
впервой половине выдоха, тормозят как инспираторные, так и экспираторные нейроны (расположены в ростральной ВЛГ).
2.Экспираторные нейроны – эфферентные нейроны центра, иннервирующие мотонейроны мышц выдоха, активны во второй половине выдоха, преимущественно при усиленном выдохе (расположены в каудальной ВЛГ).
3.Преинспираторные нейроны – интернейроны, блокирую-
щие возбуждение экспираторных нейронов, способствуют смене выдоха на вдох.
Периодическая деятельность (автоматия) дыхательного центра
Автоматия дыхательного центра – способность его обес-
печить смену вдоха и выдоха за счет своих внутренних механизмов при постоянной импульсации с хеморецепторов. В основе автоматии дыхательного центра лежит циркуляция возбуждения в его нейронах, обеспечивающая саморегуляцию вдоха и выдоха и находящаяся под выраженным произвольным корковым контролем.
Продолговатый мозг способен самопроизвольно генерировать электрические импульсы. Основная часть нейронов дыхательного центра в продолговатом мозге относится к ретикулярной формации, а эти нейроны обладают спонтанной активностью. Кроме спонтанной активности, автоматии дыхательного центра способствуют гуморальные влияния (главным образом СО2 в крови), оказываемые непосредственно на клетки центра, афферентная импульсация от хемо- и механорецепторов рефлексогенных зон, реципрокные отношения нейронов дыхательного центра, возбуждающие влияние вышележащих отделов ЦНС.
Происхождение центрального дыхательного ритма. Рит-
мическое чередование вдоха и выдоха связано с попеременными
50
разрядами инспираторных и экспираторных нейронов (рис. 17). Смена вдоха и выдоха (постоянное их чередование) обеспечивается циркуляцией возбуждения в дыхательных нейронах продолговатого мозга – главной части дыхательного центра, а также взаимодействием импульсации нейронов продолговатого мозга с импульсацией дыхательных нейронов моста и рефлексогенных зон, главной из которых является легочная (механорецепторы) (рис. 18).
а |
|
б |
|
|
|
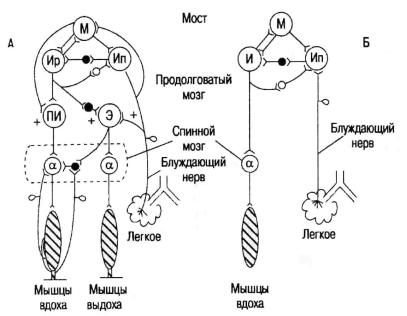
Рис. 18. Основные процессы саморегуляции вдоха и выдоха:
а– более полный вариант; б – упрощенный вариант. Нейроны:
М– ретикулярной формации моста. Ир, Ип, И, ПИ, Э – ретикулярной формации продолговатого мозга: Ир – инспираторные ранние,
Ип – инспираторные поздние, И – инспираторные (их совокупность, кроме Ип), ПИ – полные инспираторные, Э – экспираторные,
α – мотонейроны спинного мозга, черные – тормозящие, светлые – возбуждающие нейроны
При этом эфферентные импульсы от инспираторных нейронов ритмично поступают по диафрагмальному и межреберным нервам к дыхательным мышцам, что ведет к их сокращению (вдох). Пре-
51
кращение импульсации в инспираторных нейронах сопровождается расслаблением дыхательной мускулатуры (выдох).
Цикл дыхания у человека состоит из вдоха, выдоха и паузы. Классификация групп дыхательных нейронов рассмотрена выше. Разные по характеру импульсации дыхательные нейроны расположены диффузно. Возбуждающее и тормозящее взаимодействие всех типов вышеуказанных нейронов обеспечивает ритмическую деятельность дыхательного центра.
Большинство экспираторных нейронов является антиинспираторными, и только часть из них посылает свои импульсы к мышцам выдоха. Они возбуждаются под влиянием афферентной импульсации блуждающих нервов и нейронов моста. Большинство инспираторных нейронов обладает непрерывной спонтанной импульсной активностью, которая преобразуется в фазную благодаря тормозным реципрокным влияниям экспираторных и поздних инспираторных нейронов. После перерезки блуждающих нервов и ствола мозга между мостом и продолговатым мозгом наблюдается длительный тетанус инспираторных мышц (инспираторное апноэ), что также свидетельствует о постоянной активности инспираторных нейронов.
Таким образом, во время возбуждения инспираторных нейронов, обеспечивающих вдох, экспираторные клетки заторможены, т.е. «молчат», и наоборот. Это позволяет предположить, что инспираторные и экспираторные клетки оказывают друг на друга реципрокное тормозное влияние, т.е. между центрами вдоха и выдоха существуют реципрокные отношения.
Однако после выхода животного из наркоза восстанавливается ритмичное дыхание, что демонстрирует высокую степень автоматии главной части дыхательного центра продолговатого мозга и компенсационные возможности ЦНС в случае ее повреждения.
Схема саморегуляции вдоха и выдоха
Каждый дыхательный цикл начинается с возбуждения ранних инспираторных нейронов дыхательного центра, расположенного в продолговатом мозге. Затем возбуждение переходит на полные инспираторные нейроны. В процессе циркуляции
52
возбуждения импульсы по возвратным связям поступают к предшествующим нейронам и тормозят их.
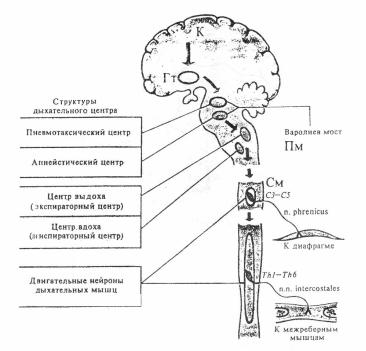
Полные инспираторные и экспираторные нейроны по нисходящим путям посылают импульсы к мотонейронам спинного мозга, иннервирующим дыхательные мышцы. Диафрагма иннервируется аксонами мотонейронов, расположенных на уровне III-IV шейных сегментов спинного мозга.
Мотонейроны, отростки которых образуют межреберные нервы, иннервирующие межреберные мышцы, расположены в передних рогах (III-ХII) грудных сегментов спинного мозга (рис. 19). Поскольку при спокойном дыхании выдох обычно осуществляется за счет потенциальной энергии, накопленной во время вдоха, экспираторные нейроны и мышцы не показаны.
В верхней части моста головного мозга, в варолиевом мосту (медиальное парабрахиальное ядро), находится пневмотаксический центр, а в нижней – апнейстический центр (рис. 19).
Рис. 19. Дыхательный центр (его компоненты) и эфферентные нервы
53
Апнейстический центр оказывает постоянную стимуляцию инспираторных нейронов, способствует вдоху и пролонгирует его. Источники дыхательных ритмов продолговатого мозга и варолиева моста связаны между собой восходящими и нисходящими нервными путями и функционируют согласованно. Так, инспираторные нейроны, возбуждаясь, посылают импульсы в пневмотаксический центр, который, в свою очередь, запускает автоматию экспираторного центра, а экспираторные нейроны тормозят инспираторные (рис. 20). Пневмотаксический центр моста тормозит инспираторные нейроны, ограничивает длительность вдоха и повышает частоту дыхания.
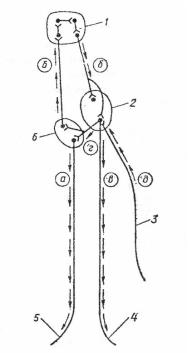
Рис. 20. Схема нервной регуляции дыхания:
а– нервные импульсы, идущие от центра вдоха; б – импульсы
кпневмотаксическому центру и к центру выдоха; в – импульсы от центра выдоха; г, д – импульсы к центрам вдоха и выдоха
от рецепторов легких; 1 – пневмотаксический центр; 2 – центр выдоха; 3 – чувствительные волокна блуждающего нерва от рецепторов растяжения легких; 4, 5 – двигательные нервные волокна к межреберным мышцам и диафрагме; 6 – центр вдоха (по К. Вилли и В. Детье, 1975)
54
Перерезка в нижней части моста приводит к удлинению вдоха, при дополнительной перерезке блуждающих нервов происходит апнейзис (глубокие длительные вдохи и короткие выдохи).
Таким образом, пневмотаксический центр контролирует деятельность нижерасположенных центров вдоха и выдоха и обеспечивает правильное чередование циклов дыхательных движений. Полагают, что он связан лишь с «тонкой настройкой» дыхательного ритма, поскольку нормальный ритм может сохраняться и в отсутствии данного центра.
РЕФЛЕКТОРНАЯ РЕГУЛЯЦИЯ ДЫХАНИЯ. МЕХАНИЗМ СМЕНЫ ДЫХАТЕЛЬНЫХ ФАЗ
Значение механорецепторов и блуждающих нервов в регуляции частоты и глубины дыхания
Дыхательный центр получает афферентные импульсы не только от хеморецепторов, но и от прессорецепторов сосудистых рефлексогенных зон, а также от механорецепторов легких (рецепторы растяжения легочной паренхимы), дыхательных путей (трахеи, бронхов, бронхиол) и дыхательных мышц (рецепторы растяжения – проприорецепторы – мышечные веретена).
Таким образом, дыхательный ритм, зарождающийся в структурах ЦНС, может изменяться под влиянием периферических импульсов. Так, если раздуть легкие, то вдох рефлекторно затормозится и начнется выдох. Напротив, если уменьшить объем легких, то произойдет глубокий вдох, что свидетельствует о том, что к дыхательным центрам постоянно поступает импульсация, сигнализирующая о степени растяжения легких, и под влиянием этой импульсации по принципу отрицательной обратной связи запускается соответствующее дыхательное движение (рис. 20, 21А). Этот механизм носит название рефлекса Геринга – Брейера (в честь авторов). Рефлекс Геринга – Брейера, являясь одним из механизмов рефлекторной саморегуляции дыхания, обеспечивает смену актов вдоха и выдоха.
55

Рис. 21. А – роль афферентных влияний блуждающего нерва в регуляции дыхания: а – кривая дыхания; б – просвет альвеол; в – импульсы, отводимые от одиночного афферентного блуждающего нерва;
г – тормозящее влияние экспираторного центра на инспираторный; д – импульсы, отводимые от инспираторного центра к дыхательной мускулатуре. Б – роль блуждающего нерва в регуляции смены фаз дыхания
При растяжении альвеол во время вдоха от рецепторов растяжения легких по афферентным блуждающим нервам импульсы идут к экспираторным нейронам, которые, возбуждаясь,
56
тормозят активность инспираторных нейронов, что приводит к пассивному выдоху. Легочные альвеолы спадаются, и нервные импульсы от рецепторов растяжения уже не поступают к экспираторным нейронам. Активность их падает, что создает условия для повышения активности инспираторной части дыхательного центра и осуществления активного вдоха. Кроме того, активность инспираторных нейронов усиливается при нарастании концентрации углекислого газа в крови после выдоха, что также способствует появлению вдоха.
При перерезке блуждающих нервов рефлекс Геринга – Брейера исчезает, и дыхание становится более медленным и глубоким (рис. 21Б). Роль эфферентного звена Геринга – Брейера играют двигательные нервы, снабжающие дыхательную мускулатуру.
Физиологическое значение рефлекса Геринга – Брейера состоит в ограничении дыхательных экскурсий. Благодаря этому достигается соответствие глубины дыхания сиюминутным условиям функционирования организма, при котором работа дыхательной системы совершается более экономично. Кроме того, в экстремальных условиях рефлекс Геринга – Брейера препятствует перерастяжению альвеол легких.
Рефлексы с межреберных мышц
К регуляции дыхательных движений относятся также социальные рефлексы с дыхательных мышц (поперечно-поло- сатая мускулатура). Эти мышцы содержат рецепторы растяжения – проприорецепторы – мышечные веретена. В случае, если либо вдох, либо выдох затруднены, мышечные веретена соответствующих мышц возбуждаются и в результате сокращения этих мышц усиливаются (проприорецептивный рефлекс). Благодаря этим особенностям межреберной мускулатуры достигается соответствие механических параметров дыхания сопротивлению дыхательной системы. Кроме того, афферентная импульсация от мышечных волокон поступает также к дыхательным центрам, обусловливая изменения деятельности дыхательной мускулатуры.
57
Защитные дыхательные рефлексы
На дыхание рефлекторно влияют раздражения слизистых оболочек дыхательных путей. Возникающие при этом чиханье и кашель являются защитными дыхательными рефлексами. Механические и химические раздражения (слизь, пыль, инородные тела) слизистой оболочки носа вызывают чиханье – глубокий сильный выдох через нос, способствующий выбрасыванию раздражителя. Афферентным путем этого рефлекса является тройничный нерв.
Кашель возникает при раздражении механо- и хеморецепторов гортани, трахеи и бронхов механическими и химическими раздражителями. Кашель и чиханье начинаются с возникающего рефлекторно глубокого вдоха. Затем происходят спазм голосовых связок, приводящий к закрытию голосовой щели, и одновременно сокращение мышц, обеспечивающее форсированный выдох. Вследствие этого давление воздуха в альвеолах, бронхах и трахее резко повышается, что приводит к раскрытию голосовой щели и к выбросу воздуха из легких толчком в верхние дыхательные пути и наружу через нос (при чиханье) или через рот (при кашле). Пыль, грязь, инородные тела увлекаются этой быстрой струей воздуха и выбрасываются из легких и дыхательных путей. К защитным рефлексам относится также опускание надгортанника при глотании.
Сильное возбуждение рецепторов гортани, возникающее, например, при внезапном вдыхании паров аммиака, хлора, сернистого ангидрида, табачного дыма и других раздражающих веществ в высоких концентрациях вызывает рефлекторный спазм голосовой щели, а иногда даже мускулатуры бронхов, сопровождающийся кратковременной рефлекторной задержкой дыхания.
Гуморальная регуляция дыхания
Гуморальная регуляция дыхания осуществляется путем изменения возбудимости дыхательного центра (ДЦ) при действии химических раздражителей и биологически активных веществ, поступающих в кровь.
58

рН артериальной крови и напряжение в ней СО2 и О2 не только непосредственно зависят от внешнего дыхания, но и оказывают влияние на вентиляцию легких.
Значение газового состава крови в регуляции дыхания было показано Л. Фредериком (1901) путем опыта с перекрестным кровообращением. Для этого у двух собак перерезали и соединили перекрестно их сонные артерии и отдельно их яремные вены. После такого соединения сосудов и зажатия других сосудов шеи голова первой собаки снабжалась кровью не от собственного туловища, а от туловища второй собаки, голова же второй собаки – от туловища первой (рис. 22).
а |
|
б |
|
|
|
Рис. 22. Опыт с перекрестным кровообращением (по Л. Фредерику) Примечание. Пережатие трахеи у собаки а вызывает одышку у собаки б, одышка собаки б вызывает замедление дыхания у собаки а
Если у одной из этих собак зажать трахею и таким образом производить удушение организма, то через некоторое время у нее происходит остановка дыхания (апноэ), у второй собаки возникает резко одышка (диспноэ). Это объясняется тем, что зажатие трахеи у первой собаки вызывает накопление СО2 в крови ее туловища (гиперкапнию) и уменьшение содержания кислорода (гипоксемию). Кровь из туловища первой собаки поступает в голову второй собаки и стимулирует ее дыхательный центр. В результате возникает усиленное дыхание (гипервентиляция) у второй собаки, что приводит к снижению напряжения СО2 и повышению О2 в крови сосуда туловища второй собаки. Богатая кислородом и бедная углекислым газом кровь из туловища этой собаки поступает в голову первой и вызывает у нее
апноэ (рис. 23).
59