

2 курс / Гистология / Мяделец.Частная гистология
.pdfфолликулы (при этом блестящая оболочка деформируется и имеет характерный вид). Одновременно сильно разрастаются интерстициальные клетки сосудистой теки. Они гипертрофируются и становятся похожими на текалютеоциты. Эти клетки интенсивно синтезируют овариальные гормоны (преимущественно эстрогены).
Физиологическая роль атрезии до конца не изучена. Поскольку при этом образуется большое количество гормонпродуцирующих интерстициальных клеток, то это обстоятельство может иметь значение для дополнительного образования гормонов, в частности, андрогенов, которые в последующем могут служить источником эстрогенов при половом созревании. Поскольку наибольшее количество атретических фолликулов образуется перед половым созреванием, то они могут служить источником эстрогенов в этот период. Кроме того, посредством атрезии организм может избавляться от лишних, а также дефектных половых клеток. Продукты их распада используются для синтетических процессов.
Причины, приводящие к атрезии фолликулов, до конца не исследованы. Предполагается, что атрезии подвергаются фолликулы, на плазмолемме которых либо отсутствуют рецепторы к гонадотропинам, либо их мало. Некоторые исследователи придают большое значение увеличенной продукции андрогенов, синтезируемых гипертрофированными интерстициальными клетками подвергающегося атрезии фолликула и инициирующих атрезию других фолликулов.
ОТЛИЧИТЕЛЬНЫЕ ОСОБЕННОСТИ ОВОГЕНЕЗА. Так же, как и сперматогенез, овогенез является проявлением генеративной функции. Он протекает аналогично сперматогенезу, но имеет некоторые особенности.
1.Фаза размножения происходит во внутриутробном периоде. Значительная часть образовавшихся в эмбриональном периоде половых клеток погибает при атрезии, а меньшая часть вступает в фазу малого роста, которая на длительное время блокируется на стадии профазы мейоза I.
2.Фаза роста делится на малый и большой рост. Малый рост протекает медленно от момента рождения до полового созревания без стимулирующего влияния гормонов. Он связан с постепенным накоплением желточных включений. Большой рост осуществляется после полового созревания, протекает быстро, при стимулирующем влиянии фоллитропина в ходе менструального цикла и связан с быстрым накоплением трофических включений. В этот период овоцит завершает профазу мейоза I. В овоците успевает произойти кроссинговер и образуются тетрады.
3фаза овогенеза - созревание - заканчивается образованием вторичного овоцита, который блокируется в метафазе второго деления мейоза. При оплодотворении в него проникает сперматозоид, и овоцит называется пенетрированным овоцитом. Проникновение сперматозоида стимулирует овоцит, и он завершает второе мейотическое деление. В результате образу-
461
ется овотида и второе редукционное тельце. В конечном итоге в процессе овогенеза формируются одна овотида и три редукционных тельца, т.е. деление клеток в мейозе неравномерное. Таким образом, при овогенезе стадия зрелой яйцеклетки как таковая отсутствует, есть стадия овотиды, которая образуется при пенетрации сперматозоидом вторичного овоцита. Оводида млекопитающих характеризуется тем, что содержит два раздельных гаплоидных набора хромосом в форме мужского и женского пронуклеусов. Соединение этих наборов в единый диплоидный ведет к образованию зиготы. При отсутствии оплодотворения половая клетка дегенерирует на стадии вторичного овоцита.
4. Фаза формирования отсутствует.
ЦИТОФИЗИОЛОГИЯ ЭНДОКРИННЫХ ФУНКЦИЙ ЯИЧНИКОВ.
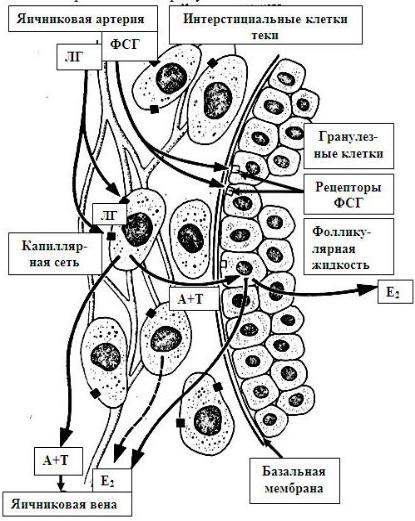
Выработка женских половых гормонов (эстрогенов). В настоящее время доминирующей является теория двухкомпонентного образования
эстрогенов (Рис. 24.6). Как считают, они образуются при содружестве двух видов клеток: фолли-
кулярных клеток и
Рис. 24.6. Схема воздействия гонадотропных гормонов на фолликул и синтез эстрогенов (по Д. Бэйрду)
ЛГ – лютеинизирующий гормон; ФСГ – фолликулостимулирующий гормон; А – андростендион; Т
– тестостерон; Е2 – эстрадиол
интерстициальных клеток -сосудистой теки. Вначале интерстициальные клетки из холестерина синтезируют мужские половые гормоны. Эти гормоны затем поступают в фолликулярные клетки, которые путем ароматиза-
ции из андрогенов синтезируют эстрогены. Часть андрогенов уходит в общую циркуляцию и оказывает действие на весь организм. Таким образом, в
462
яичнике образуются и женские, и мужские половые гормоны.
Кроме интерстициальных клеток теки фолликула, андрогены синтезируются так называемыми гилусными клетками - эндокринными клетками, схожими с клетками Лейдига яичка и локализующимися в области ворот яичника (гилуса). Количество этих клеток увеличивается при беременности и в менопаузу. Гиперплазия гилюсных клеток может приводить к маскулинизации. Биосинтез половых гормонов, имеющих стероидную природу, происходит в гладкой эндоплазматической сети и митохондриях клетокпродуцентов.
2. Желтое тело продуцирует два гормона беременности: стероидный гормон прогестерон и гормон белковой природы релаксин. Кроме них, в желтом теле синтезируются эстрогены, в небольшом количестве оксито-
цин, андрогены и простагландины.
4. Клетки зернистого фолликулярного эпителия образуют гормон ин-
гибин, а также фактор, аналогичный гонадолиберинам и, возможно, фол-
литропин.
ВАСКУЛЯРИЗАЦИЯ, ИННЕРВАЦИЯ И РЕГЕНЕРАЦИЯ ЯИЧНИКОВ. Кровоснабжение яичника осуществляется из яичниковой артерии, которая после вхождения в орган через его ворота приобретает сильно извитой ход. В корковом веществе артерии распадаются на большое количество мелких радиальных ветвей и далее до капилляров, образующих густые сети вокруг фолликулов. Следует отметить, что в период роста фолликулов и при формировании желтого тела сосудистый аппарат яичника проявляет высокую способность к функциональной перестройке: во внутренней теке фолликула и затем в образующемся желтом теле имеет место так называемый функциональный ангиогенез, или новообразование сосудов. Напротив, при обратном развитии желтого тела сосудистое русло редуцируется.
Капилляры сливаются в вены, также имеющие спиральный ход и анастомозирующие между собой. Вены образуют два сплетения: в мозговом веществе и в области ворот. Лимфокапилляры берут свое начало в окружности фолликулов. Затем они сливаются в более крупные лимфососуды коркового вещества, которые идут в мозговое вещество и к воротам. Только после выхода из ворот в лимфососудах появляются клапаны.
Яичник получает как чувствительную, так и эфферентную иннервацию. Афферентные пути образованы дендритами псевдоуниполярных нейронов нижнегрудных, верхнепоясничных и нижнекрестцовых сегментов спинного мозга. В яичнике имеются как свободные, так и несвободные нервные окончания (в т.ч. тельца Фатер-Пачини).
Эфферентная иннервация (в основном сосудов) обеспечивается как симпатической, так и парасимпатической нервной системой. Центры симпатической нервной системы находятся в боковых промежуточных ядрах боковых рогов серого вещества нижнегрудных и верхнепоясничных сегментов
463
спинного мозга. Отростки нейронов этих ядер идут к симпатическому стволу и затем к ганглиям яичникового и маточного сплетений. Постганглионарные волокна направляются к сосудам яичника. Парасимпатические преганглионарные волокна образованы отростками клеток боковых промежуточных ядер промежуточного столба 2-4-го крестцовых сегментов спинного мозга. По передним корешкам они вступают в соответствующие спинномозговые нервы и доходят до параорганных сплетений, где заканчиваются синапсами на клетках Догеля I типа. Аксоны этих нейронов иннервируют сосуды. И симпатические, и парасимпатические нервные волокна образуют обильные нервные сети вокруг фолликулов, желтых тел и в мозговом веществе. Возбуждение симпатической нервной системы вызывает сокращение, а парасимпатической - расслабление гладкой мускулатуры сосудов яичника. Нервные влияния на фолликулогенез и лютеогенез незначительны.
Физиологическая регенерация яичников осуществляется на клеточном уровне и связана периодическим ростом фолликулов. Входящие в их состав фолликулярные клетки обладают высокой способностью к делению. Такой же высокой регенераторной способностью обладает покровный эпителий яичника, содержащий мозаично распределенные по пласту камбиальные клетки. Соединительная ткань восстанавливается за счет собственных камбиальных клеток. Как важную особенность физиологической регенерации яичника следует отметить упоминавшийся ранее функциональный ангиогенез - новообразование сосудов при росте фолликулов и формировании желтого тела.
Репаративная регенерация яичников также осуществляется на клеточном уровне, причем ее объем зависит от количества удаленной паренхимы. При удалении 4/5 части одного из яичников наблюдается атрофия остатка органа и лишь изредка отмечаются признаки гипертрофии. Однако в клинической практике описаны случаи регенерации одного из двух удаленных путем полного удаления (вылущивания, энуклеации) яичников. Предполагается, что в этих случаях регенерация происходит за счет незначительных остатков паренхимы, сохранившихся на капсуле.
Удаление половины одного из яичников при сохранении второго также ведет к атрофии остатка резецированного органа. Одновременно наблюдается незначительное увеличение веса и размеров интактного яичника, гистологически - заметное нарастание количества желтых тел, что свидетельствует о компенсаторном увеличении частоты в этом яичнике овуляций. Удаление половины одного яичника и полного второго яичника ведет к резкой гипертрофии остатка резецированного органа. Его вес возрастает в 3 раза. На раневой поверхности не формируется рубец, она эпителизируется за счет покровного мезотелия. Возрастание размеров остатка органа происходит за счет усиленного созревания фолликулов и образования большого количества желтых тел.
464
РЕГУЛЯЦИЯ ГЕНЕРАТИВНОЙ И ЭНДОКРИНЫХ ФУНКЦИЙ ЯИЧНИКОВ. Существует несколько уровней регуляции функций яичника.
1.Внутрисистемный уровень. Эстрогены регулируют рост фолликулов; прогестерон подавляет их рост и синтез эстрогенов. Ингибин предотвращает в развивающемся фолликуле преждевременное вступление в мейоз овоцита. Для внутрисистемной регуляции функций яичника определенное значение имеют также секретируемые в яичнике простагландины и биологически активные вещества.
2.Гипофизарный уровень регуляции. В гипофизе вырабатывается два гонадотропных гормона: фоллитропин и лютропин. Рост фолликулов в яичнике и выработка эстрогенов стимулируется фоллитропином. Однако, как отмечалось, для выработки эстрагенов необходимо содружество двух типов клеток: интерстициальных и фолликулярных. Стимуляция интерстициальных клеток выполняется лютропином. Поэтому для активации образования эстрагенов необходимо взаимодействие этих двух гормонов. Лютропин стимулирует овуляцию и образование желтого тела, продукцию им прогестерона. Для развития желтого тела необходим также гормон аденогипофиза лактотропин (маммотропин), который участвует в формировании желтого тела и усиливает продукцию им прогестерона.
3.Гипоталамический уровень. В гипоталамусе вырабатывается два типа гормонов: гонадолиберины и гонадостатины. Гонадолиберины стимулируют выработку гипофизом гонадотропинов. Гонадостатины оказывают на гипофиз противоположный эффект. В отличие от мужского организма,
секреция гипоталамических гормонов идет циклически. Это обеспечивает цикличность изменений в женской половой системе.
4.Лимбический уровень. Лимбическая система стимулирует выделение гипоталамусом гонадолиберинов.
5.Эпифизарный уровень: гормоны эпифиза тормозят выработку гонадотропных гормонов гипофизом.
6.Корковый уровень регуляции.
ФУНКЦИОНАЛЬНАЯ МОРФОЛОГИЯ МАТОЧНЫХ ТРУБ
Маточные трубы - парные органы, которые связывают полость брюшины с полостью матки и обеспечивают захват, продвижение неоплодотворенной яйцеклетки в сторону матки, служат местом ее оплодотворения и начальных этапов эмбриогенеза.
РАЗВИТИЕ. Источником развития маточных труб являются верхние части парамезонефральных протоков. Мезотелий серозной оболочки образуется из мезодермы висцерального листка спланхнотома.
СТРОЕНИЕ. Анатомически маточные трубы состоят из воронковой,
ампулярной, истмической и маточной частей. Они являются органами слоистого типа (Рис. 24.7). Стенка маточных труб (за исключением маточ-
465
ной части) образована тремя оболочками: слизистой, мышечной и серозной (в маточной части серозная оболочка отсутствует).
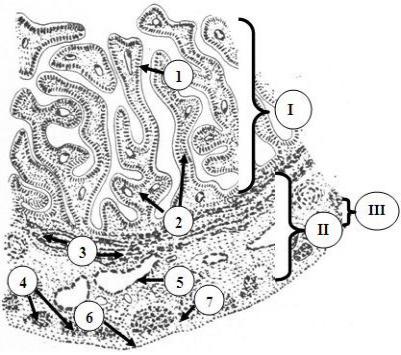
Рис. 24.7. Строение маточной трубы
I – слизистая оболочка, формирующая складки: 1
– однослойный столбчатый эпителий; 2 – собственная пластинка; II – мышечная оболочка: 3 – циркулярный, 4 – продольный слои; 5 – кровеносные сосуды; III – серозная оболочка: 6 – мезотелий; 7 – собственная пластинка серозной оболочки
Слизистая оболочка состоит из двух слоев: эпителиального и собственной пластинки. В воронковой и ампулярной частях она формирует многочисленные сложные разветвленные продольные складки. При приближении к матке число складок и их размеры постепенно уменьшаются, и, в конце концов, они превращаются в невысокие гребешки. Эпителиальный слой представлен однослойным призматическим эпителием. В его составе имеются два вида клеток: реснитчатые и секреторные. В последнее время в эпителии описывают вставочные и базальные клетки. Реснички реснитчатых клеток совершают колебательные движения в сторону полости матки, что обеспечивает продвижение в этом направлении оплодотворенной яйцеклетки. Число реснитчатых клеток увеличено на фимбриях маточных труб, т.е. в той их части, которая открывается в полость брюшины. Количество ресничек убывает от воронки к истмическому отделу. Железистые клетки вырабатывают слизь, содержащую питательные вещества для яйцеклетки, а также вещества, активирующие сперматозоиды перед оплодотворением (капацитация) и ферменты, участвующие в денудации яйцеклетки (см. функции маточных труб). Секреция различных по локализации эпителиоцитов осуществляется либо по мерокриновому, либо по апокриновому типу. Эпителий секретирует также простагландины, осуществляющие местную паракринную регуляцию функций маточных труб.
В средней части соотношение слизистых и мерцательных клеток равно 10:1. В эпителии встречаются внутриэпителиальные лимфоциты. Вместо однослойного могут появляться участки многослойного плоского эпителия,
466
что связано с эндокринными нарушениями. Предполагается также дистопия в яйцевод многослойного эпителия влагалища. Cреди эпителиоцитов имеются менее дифференцированные клетки, за счет которых происходит физиологическая регенерация эпителия.
В собственной пластинке, образованной РСТ, встречаются крупные полигональные, богатые гликогеном децидуальные клетки (строение и функции децидуальных клеток описаны в разделе “Эмбриология человека”).
Слизистая оболочка маточных труб претерпевает циклические изменения в менструальном цикле. Перед овуляцией формируемые ею складки продольные, а в лютеиновую фазу становятся сложнорельефными. Высота эпителия и его секреторная активность максимальны в фазе секреции и минимальны в фазе пролиферации. Движения ресничек наиболее выражены перед овуляцией, сразу после ее наступления и во время ее. Слизистая оболочка воронковой части маточных труб перед овуляцией набухает из-за переполнения кровью вен. Это способствует охвату бахромкой воронки яичника и приему яйцеклетки. Подобные трансформации слизистой оболочки маточных труб опосредуются женскими половыми гормонами.
Мышечная оболочка состоит из внутреннего циркулярного и наружного продольного слоев гладкой мышечной ткани. Циркулярный слой выражен сильнее. В истмическом и маточном отделах мышечная оболочка дополняется третьим слоем - внутренним продольным, который обеспечивает расширение просвета яйцеводов для свободного прохождения развивающегося зародыша. Перистальтические сокращения мышечной оболочки обусловливают передвижение яйцеклетки и зародыша по яйцеводу к матке. Они активируются эстрогенами и подавляются прогестероном. Наружная оболочка - серозная, состоит из мезотелия и слоя РСТ. Последний имеет значительную толщину. В маточной части она отсутствует.
ФУНКЦИИ МАТОЧНЫХ ТРУБ. 1. Захват яйцеклетки и обеспечение ее передвижения в направлении полости матки. В момент овуляции сосуды фимбрий переполняются кровью, фимбрии увеличиваются в объеме в виде воронки охватывают яичник. Передвижение яйцеклетки или зародыша осуществляется за счет биения ресничек и перистальтических движений, вызванных сокращением мышечной оболочки.
2. Секреторная функция заключается в выработке слизи. Слизь способствует передвижению яйцеклетки, содержит питательные вещества для яйцеклетки и зародыша (гистотрофный тип питания), а также вещества, активирующие движение сперматозоидов (капацитация). В слизи содержатся ферменты, которые осуществляют освобождение яйцеклетки от фолликулярных клеток - денудацию. Денудация необходима для последующего оплодотворения, поскольку при этом обнажается блестящая оболочка, несущая рецепторы для сперматозоидов (ZP2, ZP3 и др.). К секреторной (эндокринной) функции яйцеводов относится также биосинтез эпителиоцитами слизи-
467
стой оболочки простагландинов.
3. Маточные трубы обеспечивают ранние этапы эмбриогенеза. В них в норме происходит оплодотворение яйцеклетки. От длины яйцеводов зависит, где в матке имплантируется зародыш. Чем длиннее яйцеводы, тем дальше в них находится зародыш и продвигается эмбриогенез. При этом более выраженными становятся адгезивные и литические свойства трофобласта, и зародыш имплантируется в эндометрий матки ближе к устью яйцеводов. Длинные яйцеводы могут стать одной из причин внематочной (трубной) беременности. Напротив, при коротких яйцеводах адгезивные свойства трофобласта к моменту попадания зародыша в полость матки оказываются недостаточными, и имплантация может произойти ближе к шейке матки. Это так называемое низкое предлежание плаценты (иногда у внутреннего зева шейки матки). При нем во время раскрытия внутреннего зева при начале родовой деятельности может наступить отслойка плаценты с развитием опасного для жизни роженицы и плода кровотечения.
КЛИНИЧЕСКИЕ АСПЕКТЫ ГИСТОФИЗИОЛОГИИ МАТОЧНЫХ ТРУБ. При воспалительных процессах маточных труб (сальпингитах), имеющих гонорейную и негонорейную природу и локализующихся в слизистой оболочке, происходит разрушение эпителия с формированием в собственной пластинке рубцовой ткани, иногда спаек. Эти рубцы и спайки могут полностью или частично закрывать просвет яйцевода. При полной обтурации просвета оплодотворение яйцеклетки невозможно - развивается бесплодие. При неполном закрытии сперматозоиды могут проникать к яйцеклетке и оплодотворять ее, однако последующая миграция зародыша в полость матки невозможна. В этом случае зародыш либо погибает, либо развивается внематочная (трубная) беременность, при которой ворсины хориона разрушают стенку яйцеводов. Это может привести к развитию опасного для жизни женщины кровотечения.
ФУНКЦИОНАЛЬНАЯ МОРФОЛОГИЯ МАТКИ
ФУНКЦИИ МАТКИ. 1. Матка является вместилищем для развивающегося плода. 2. Матка активно участвует в процессе родов благодаря сократительной деятельности миометрия. Кроме того, шейка матки вместе с влагалищем формирует родовой канал, по которому продвигается рождающийся плод. 3. Секреторная функция матки заключается в выработке слизистого секрета. 4. Участие в образовании плаценты и плодных оболочек: эндометрий матки формирует материнскую часть плаценты, а также децидуальную капсулярную оболочку. 5. Эндокринная функция заключается в выработке простагландинов, релаксина, андрогенов. Простагландины и релаксин синтезируются маточными железами, а половые гормоны - интерстициальными клетками, расположенными в РВНСТ, располагающейся между слоями мышечной оболочки. 6. Участие в формировании женского оргазма.
ИСТОЧНИКИ РАЗВИТИЯ. Матка развивается из нижних частей пара-
468
мезонефральных протоков в результате их слияния (см. выше). В результате этого тело матки плода имеет слабо развитую двурогость, которая исчезает к 4-му месяцу эмбриогенеза. Мезотелий периметрия образуется из висцерального листка спланхнотома.
СТРОЕНИЕ (Рис. 24.8). В анатомическом отношении в матке выделяют
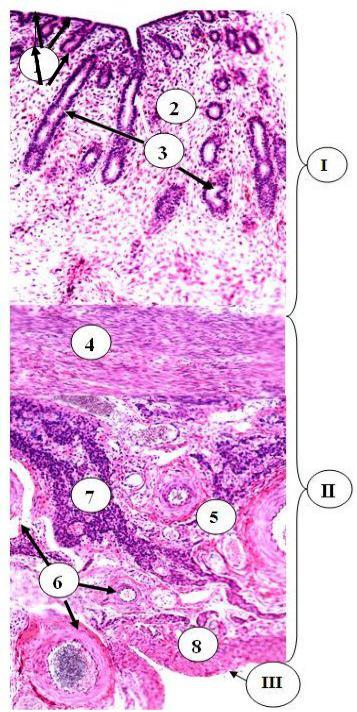
Рис. 24.8. Строение тела матки I – слизистая оболочка тела матки (эндометрий): 1 – однослойный столбчатый эпителий; 2 – собственная пластинка (строма) эндометрия; 3 – маточные железы;
II - мышечная оболочка тела матки (миометрия): 4 – подслизистый, 5 – сосудистый слои; 6 – кровеносные сосуды сосудистого слоя миометрия; 7 – скопления интерстициальных клеток; 8 – надсосудистый слой миометрия; III – серозная оболочка с мезотелием и соединительнотканной основой
тело и шейку. С гистологической точки зрения матка - орган слоистого типа, состоящий из трех оболочек: слизистой,
мышечной и серозной (адвентициальной). Эти оболоч-
ки имеют специфические названия: слизистая называется
эндометрием, мышечная - миометрием, серозная - пе-
риметрием. Так как миометрий имеет большую толщину, в нем можно выделить паренхиму (совокупность миоцитов,
формирующих слои), и строму, образованную прослойками РСТ. Поэтому матку иногда относят и к органам смешанного типа, сочетающим признаки и слоистого, и паренхиматозного органов.
Периметрий образован мезотелием и пластинкой РСТ, которая тесно сообщается с межмышечной РСТ миометрия. Он окружает матку на
469
большем протяжении. Непокрытыми остаются передняя и боковые поверхности надвлагалищной части шейки матки. В периметрии залегают симпатические ганглии и сплетения. В тех участках, где периметрий отсутствует, наружной оболочкой является адвентициальная оболочка. Здесь вокруг шейки матки имеется скопление жировой ткани, которое называется параметрием. Он имеет неодинаковую толщину в разных участках. В параметрии находятся крупные кровеносные, лимфатические сосуды и нервные стволы, входящие в матку. Иногда здесь могут наблюдаться воспалительные процессы, которые называются параметритом. Обычно это происходит в первые дни после осложненных родов. Миометрий сильно развит, имеет большую толщину - до 1,5 см. При беременности его толщина резко возрастает. Гладкие миоциты имеют отростчатую форму и длину до 50 мкм, а при беременности увеличиваются примерно в 10-15 раз и достигают размера до
800 мкм.
Миометрий образован тремя слоями гладкой мышечной ткани. Внутренний слой называется подслизистым и имеет косое направление миоцитов. Средний слой, сосудистый, циркулярный. В нем содержатся многочисленные крупные кровеносные сосуды, причем артерии здесь имеют спиралевидный ход. Это имеет большое значение для остановки послеродовых кровотечений: сокращение среднего слоя миометрия ведет к пережатию сосудов и остановке кровотечения. Сокращение миометрия активируется гормоном окситоцином, который выделяется в это время в большом количестве из задней доли гипофиза. При затяжных родах в результате развития усталости миометрия его сокращение недостаточное или не происходит совсем. В этом случае возникает угроза смерти женщины от кровопотери. Эффективным кровеостанавливающим средством в этом случае является окситоцин или его синтетические аналоги. Иногда эти вещества оказываются неэффективными, и тогда для спасения жизни женщины прибегают к надвлагалищной ампутации матки.
Наружный слой миометрия - надсосудистый - имеет косое направление гладких миоцитов, но их ход противоположен таковому во внутреннем слое. Указанные слои миометрия отделяются друг от друга неотчетливо, поскольку пучки миоцитов переходят из одного слоя в другой и анатомически связывают их в единое мощное функциональное образование, эффективно обеспечивающее роды. Между слоями миометрия находится РСТ, в которой встречаются скопления крупных эпителиоидных интерстициальных клеток, продуцентов андрогенов.
Некоторые авторы выделяют четвертый, субсерозный мышечный слой.
Эндометрий неподвижно соединяется с миометрием, т.к. подслизистая оболочка отсутствует. Поэтому его складчатость не выражена. С гистологической точки зрения эндометрий состоит из двух слоев: эпителия и собст-
470