

2 курс / Гистология / Мяделец.Частная гистология
.pdfГТБ резко уменьшает концентрацию антигенов, поступающих к тимоцитам коры. При нарушении барьера в тимусе могут возникать лимфоидные узелки и образовываться плазмоциты, т.е. начинается антигензависимый лимфопоэз.
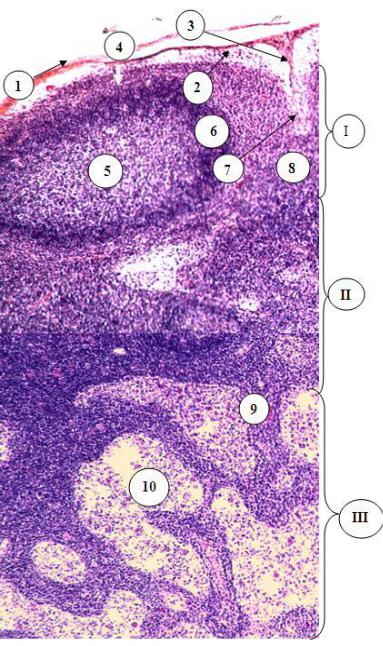
Рис. 21.8. Строение тимуса А – малое, Б – большое увеличение (мозговое вещество):
1 – соединительнотканная капсула; 2 – корковое вещество: 3
– соединительнотканные септы, 4 – ядра ретикулярных эпителиоцитов; 5 – мозговое вещество: 6 – тимусные тельца; 7 - ретикулярный эпителиоцит мозгового вещества; 8 – ретикулярный эпителиоцит тимусного тельца
МОЗГОВОЕ ВЕЩЕСТВО в отличие от коркового помимо ретикулярной эпителиальной стромы содержит соединительнотканную строму, а также лимфоциты, количество которых значительно меньше, чем в корковом веществе (3-5% от всех лимфоцитов тимуса). Часть лимфоцитов мигрирует сюда из коркового вещества, чтобы на границе с корой через посткапиллярные венулы покинуть тимус. Другая часть лимфоцитов мозгового вещества, как полагают, является лимфоцитами, поступившими из периферических органов иммуногенеза. Их роль не совсем ясна, но предполагают, что они могут приносить с периферии в тимус определенную информацию, которая учиты-
361
вается при образовании новых лимфоцитов в корковом веществе, т.е. играют регуляторную функцию.
В мозговом веществе отсутствует гемато-тимический барьер. В нем со-
держатся эпителиальные тимусные тельца Гассаля. Они образованы на-
слоенными друг на друга ретикулярными эпителиоцитами (РЭ VI типа). Размеры телец Гассаля и их численность увеличиваются с возрастом и (особенно резко) - при стресс-воздействиях. С тельцами Гассаля часто взаимодействуют макрофаги и лимфоциты. В периферических ретикулоэпителиоцитах телец Гассаля обнаруживаются признаки секреторной активности: имеются развитый синтетический и секреторный аппараты, гранулы секрета. Функциональное значение телец Гассаля до конца не выяснено. Возможными их функциями являются:
1.Образование тимических гормонов (эндокринная функция).
2.Место окончательного разрушения аутореактивных Т-лимфоцитов. Возможно, ретикулоэпителиоциты телец Гассаля имеют отношение к запуску программы апоптоза и к заключительным этапам элиминации аутоагрессивных Т-лимфоцитов.
ВОЗРАСТНАЯ И АКЦИДЕНТАЛЬНАЯ ИНВОЛЮЦИЯ ТИМУСА. После 20 лет происходит уменьшение размеров тимуса, его обратное развитие (возрастная инволюция). В органе уменьшается число лимфоцитов, лимфоидная ткань замещается жировой (Рис. 21.9). Половые гормоны усиливают и ускоряют инволюцию тимуса, тогда как кастрация вызывает его гипертрофию. В некоторых случаях нормальный процесс инволюции тимуса нарушается и орган, наоборот, увеличивается в размерах. Это состояние называется тимолимфатическим статусом. Оно сопровождается снижением устойчивости к инфекциям и токсинам и может закончиться внезапной смертью.
При стрессе под действием резко возросшего в крови содержания кортикостероидов в корковом веществе тимуса происходит массивная гибель средних и малых лимфоцитов (кортизончувствительных лимфоцитов).
Кроме того, часть лимфоцитов выбрасывается из тимуса в кровь. В результате этих процессов также наступает инволюция тимуса, которая в отличие от возрастной называется акцидентальной, или временной. После окончания действия стресс-факторов структура тимуса, в отличие от возрастной инволюции, вновь восстанавливается.
ВАСКУЛЯРИЗАЦИЯ ТИМУСА. Поступающие в тимус артерии ветвят-
ся на междольковые, внутридольковые, а затем дуговые сосуды. Дуговые артерии распадаются до капилляров, образующих глубокую сеть в коре. Меньшая часть корковых капилляров на границе с мозговым веществом пе-
реходит в посткапиллярные вены с высоким эндотелием. Через них осу-
ществляется рециркуляция лимфоцитов. Большая часть капилляров не захо-
362

дит в посткапиллярные венулы с высоким эндотелием, а продолжается в субкапсулярные венулы. Венулы переходят в выносящие вены.
В тимусе отсутствуют приносящие лимфатические сосуды. Имеются ИННЕРВАЦИЯ ТИМУСА. Тимус получает богатую иннервацию. Нерв-
ный аппарат органа связан прежде всего с сосудами. Сосудистые нервы являются как чувствительными, так и симпатическими эфферентными, оказывающими вазоконстрикторный эффект. Помимо сосудистых нервов, имеются нервные волокна, оканчивающиеся в паренхиме органа синапсами открытого типа.
Рис. 21.9. Возрастная инволюция тимуса 1 – корковое вещество; 2 – тимус-
ные тельца Гассаля; 3 – кровеносный сосуд; 4 – жировая ткань
РЕГЕНЕРАЦИЯ ТИМУСА. Физиологическая регенерация вилочковой железы осуществляется на достаточно высоком уровне, хотя и несколько более низком, чем физиологическая регенерация костного мозга. Митотический индекс клеток в корковом веществе составляет 4%, в мозговом - 2,5%. Восстановле-
ние органа после акцидентальной инволюции начинается достаточно быстро, спустя 3-4 суток после ее наступления, и сопровождается возрастанием митотического индекса в 3-4 раза. Следует подчеркнуть полное и быстрое восстановление тимуса после акцидентальной инволюции (реституция). В восстановлении органа существенную роль играет заселение его лимфоцитами из других лимфоидных органов, прежде всего из костного мозга. Иная картина отмечается при потере части органа (части доли или одной доли) после оперативных вмешательств. В этом случае тимус не способен ни к регенерации, ни к компенсаторной гипертрофии. Это связано с тем, что для полноценной регенерации органа обязательным условием является сохранение его ретикулоэпителиальной стромы.
ВТОРИЧНЫЕ ОРГАНЫ ЛИМФОИДНОЙ СИСТЕМЫ
Во вторичных (ранее - периферических) органах лимфоидной системы происходит встреча иммунокомпетентных клеток с антигенами. После этого включаются иммунные реакции, в основе которых лежит антигензависимая дифференцировка лимфоцитов. В результате образуются эффекторные клетки, инактивирующие антигены. Если уничтожение антигена идет в ре-
363
акциях клеточного иммунитета, то эффекторными клетками являются цито-
токсические Т-лимфоциты (Т-киллеры), Т-х1, NK-клетки, К-клетки и
другие клетки (см. выше). В гуморальном иммунном ответе эффекторными клетками являются плазматические клетки (плазмоциты), образующие антитела.
Все вторичные лимфоидные органы подразделяются на лимфоретику-
лярные (лимфатические узлы и селезенка) и лимфоретикулоэпителиаль-
ные (миндалины, аппендикс, лимфоидная ткань, ассоциированная с кишкой, или групповые лимфоидные узелки, пейеровы бляшки). Это подразделение учитывает формирующееся в органах микроокружение для дифференцирующихся лимфоцитов. Одиночные и групповые лимфоидные узелки слизистых оболочек не являются органами в строгом понимании, это иммунокомпетентные структуры в составе неиммунных органов, играющие большую роль в целостной иммунной системе.
В распределении периферических органов и структур (иммунокомпетентных частей в неиммунных органах) прослеживается четкий иерархический принцип. Пейеровы бляшки и одиночные лимфоидные узелки слизистых оболочек уничтожают антигены, нарушившие эпителиальный барьер и проникшие в собственную пластинку (при этом они могут растворяться в тканевой жидкости и в таком виде взаимодействовать с иммунокомпетентными клетками узелков). Если антигены прорывают эти две первые линии защиты, они с лимфой проникают в регионарные лимфоузлы и там уничтожаются. Регионарные лимфоузлы чаще располагаются в виде комплексов, и антигены при их большом количестве могут последовательно уничтожаться в нескольких лимфоузлах. Если этого не происходит, то, попадая с лимфой в кровоток, антигены заносятся в селезенку, где завершается их уничтожение. В ослабленном организме этого не происходит, и это грозит развитием септических состояний.
ЛИМФАТИЧЕСКИЕ УЗЛЫ
Лимфатические узлы (лимфоузлы) располагаются по ходу лимфососудов и уничтожают антигены, находящиеся в лимфе.
ФУНКЦИИ. 1. Кроветворная функция. У взрослого человека заключается в антигензависимой дифференцировке лимфоцитов. В эмбриональном периоде лимфоузлы в течение короткого времени выполняют функцию универсальных кроветворных органов, однако она быстро теряется. При патологии кроветворения эта функция может возобновляться у взрослых.
2. Барьерно-защитная функция: а) неспецифическая защита от антигенов заключается в фагоцитозе их из лимфы многочисленными макрофагами и “береговыми” клетками (эндотелием лимфатических синусов); б) специфическая защитная функция состоит в осуществлении специфических иммунных реакций, направленных против антигенов.
364
3.Дренажная функция. Лимфоузлы собирают лимфу из приносящих сосудов, идущих от тканей. Лимфа образуется постоянно, источником ее образования является тканевая жидкость. После прохождения через лимфоузлы лимфа очищается и в последующем поступает в венозную систему. При нарушении дренажной функции лимфоузлов наблюдается скопление тканевой жидкости в тканях - периферический отек (лимфэдема). Это явление достаточно часто встречается в клинике. Например, при мастэктомии по поводу рака молочной железы вместе с пораженным органом удаляются регионарные лимфоузлы. При этом из-за их отсутствия возникает отек верхней конечности. При закупорке лимфососудов нижней конечности филяриями (паразитическими нематодами) развивается лимфэдема нижних конечностей (слоновость, или элефантиаз).
4.Функция депонирования лимфы. В норме определенное количество лимфы задерживается в лимфоузлах и выключается из лимфотока. При необходимости она вновь поступает в лимфоциркуляцию.
5.Обменная функция - участие в обмене веществ: белков, жиров, углеводов и др. Находящиеся в лимфе питательные вещества способны захватываться макрофагами лимфоузла и подвергаться расщеплению под действием лизосомальных ферментов.
РАЗВИТИЕ. Закладка и дифференцировка лимфоузлов в эмбриогенезе происходит асинхронно. Зачатки первых лимфоузлов появляются в конце 2- го - начале 3-го месяца эмбриогенеза в виде скопления мезенхимы по ходу формирующихся лимфососудов. Мезенхима превращается в ретикулярную ткань. К концу 4-го месяца в закладку лимфоузла вселяются лимфоциты и формируются лимфоидные узелки (фолликулы). Примерно в это же время начинают формироваться синусы лимфоузла в виде эндотелиальных трубок. Лимфоузел разделяется на корковое и мозговое вещество. Однако большая часть лимфоузлов начинает формироваться во второй половине беременности.
СТРОЕНИЕ (Рис. 21.10). Общее число лимфоузлов равно примерно 1000, что составляет около 1% от массы тела. Их размеры в среднем равны
0.5- 1 см. Лимфоузлы имеют округлую почковидную форму и лежат регионарно по отношению к органам, чаще группами. С выпуклой поверхности лимфоузла в него входят приносящие лимфососуды, а с противоположной стороны, которая называется воротами, выходят выносящие лимфососуды. Кроме того, через ворота в лимфоузел проникают артерия и нервы и выходят вены.
Лимфоузлы являются паренхиматозными зональными органами, состоящими из коркового и мозгового вещества. Стромой их является капсула, отходящие от нее трабекулы и находящаяся между ними ретикулярная ткань. Паренхиму составляет совокупность гемопоэтических клеток (лимфо-
365
цитов и плазмоцитов на разных стадиях развития). В лимфоузлах можно выделить следующие структурно-функциональные компоненты.
1.Капсула, покрывающая лимфоузел снаружи и состоящая из РВНСТ с большим количеством коллагеновых волокон. В капсуле встречаются гладкие миоциты, способствующие активному продвижению лимфы, а также нервные стволики и чувствительные окончания, в том числе и инкапсулированные - пластинчатые тельца Фатер-Пачини. Болезненность лимфоузлов при воспалительных заболеваниях обусловлена отеком и сдавлением телец Фатер-Пачини. В капсуле содержатся также кровеносные сосуды: артерии, вены и гемокапилляры.
2.Трабекулы, отходящие от капсулы и образованные РВНСТ. Анастомозируя друг с другом, они формируют каркас лимфоузла. Трабекулы со-
держат единичные гладкие миоциты.
Рис. 21.10. Строение лимфатического узла
I - корковое вещество; II – паракортикальная область; III – мозговое вещество
1 – капсула; 2 – субкапсулярный синус; 3 – корковая трабекула; 4 – вторичный лимфоидный узелок; 5 – его центр размножения; 6 – мантийная зона; 7 – межузелковый синус; 8 – межузелковая часть; 9 – мозговой тяж; 10 – мозговой лимфатический синус
3. Ретикулярная ткань, заполняющая все пространство между капсулой и трабекулами. Она относится к соединительным тканям со специальными свойствами и создает микроокружение для гемопоэтических клеток.
4. В лимфоузле различают две зоны: перифери-
ческую - корковое веще-
ство - и центральную -
366
мозговое вещество.
5.Между корковым и мозговым веществом находится паракортикальная зона, которую часто не выделяют как самостоятельную зону и опреде-
ляют как глубокую кору.
6.Синусы - совокупность лимфососудов, по которым движется лимфа. Последовательность прохождения лимфы через лимфоузел и расположе-
ния синусов такова: приносящие лимфососуды краевой, или субкапсу-
лярный синус промежуточные корковые синусы промежуточные мозговые синусы воротный синус выносящий лимфососуд в облас-
ти ворот. Поскольку приносящих сосудов больше, чем выносящих, то лимфатические синусы сходятся к воротам, формируя подобие воронки, через которую лимфа проходит под давлением. Это способствует ее фильтрации. При прохождении лимфы по лимфатическим синусам осуществляется фагоцитоз содержащихся в ней антигенов макрофагами синусов.
КОРКОВОЕ ВЕЩЕСТВО. В нем обнаруживается скопление лимфоид-
ной ткани, представленной лимфоидными узелками и межузелковой ча-
стью. Лимфоидные узелки - округлые образования величиной до 1 мм.
Различают первичные, без реактивного центра, и вторичные лимфоидные узелки, имеющие реактивный центр (син. центр размножения, светлый центр).
Первичные узелки состоят в основном из малых “девственных” В- лимфоцитов, связанных с ретикулярными и фолликулярными дендритными клетками. Здесь находятся также в небольшом количестве Т-лимфоциты и макрофаги. Первичные лимфоидные узелки в наибольшем количестве встречаются во внутриутробном периоде. После рождения в связи с имеющей место постоянной антигенной стимуляцией происходит их постоянное превращение во вторичные узелки. При попадании антигена или В- лимфоцита памяти происходит бласттрансформация «девственных» В- лимфоцитов, что ведет к появлению центра размножения и формированию вторичных узелков. Они состоят из центра размножения, в котором происходит размножение В-лимфоцитов, и короны, или мантии, на периферии. Корона образована малыми В-лимфоцитами памяти, а также малыми “наивными” лимфоцитами костномозгового происхождения. Здесь же встречаются макрофаги, лимфобласты и ретикулярные клетки. Корона ассиметрична. На полюсе, обращенном в сторону мозгового вещества, она истончена, а на периферическом полюсе утолщена и формирует подобие шапочки. Латеральные поверхности короны, граничащие со светлой зоной центра размножения (см. ниже), служат местом выхода из фолликула незрелых плазмоцитов.
Реактивный центр на высоте иммунной реакции подразделяется на
темную и светлую зоны. Светлая зона состоит из светлой базальной и светлой апикальной зон. Темная зона обращена к паракортикальной зоне.
367
Здесь делятся митозом первичные В-бласты, содержащие на своей поверхности рецепторные иммуноглобулины. Они образуются из В- лимфоцитов памяти, которые поступают сюда из крови через посткапиллярные венулы с высоким эндотелием. На 4-е сутки первичные бласты превращаются в центробласты, лишенные поверхностных иммуноглобулинов. Из центробластов образуются центроциты, которые перемещаются в светлую базальную зону, где вновь начинают экспрессировать поверхностные иммуноглобулины. В это время происходит переключение класса иммуноглобулинов. В базальной светлой зоне происходит и апоптоз В-лимфоцитов, здесь же обнаруживаются содержащие апоптозные гипербазофильные тела макрофаги, а также ФДК. Центроциты начинают тесно взаимодействовать с ФДК с участием собственных рецепторов и экспрессируемых на ФДК рецепторов. Это приводит к образованию так называемых вторичных В- клеток-бластов. Они перемещаются в апикальную светлую зону центра размножения, а затем покидают его и превращаются либо в В-лимфоциты памяти, либо в плазмоциты. Если центроцит не провзаимодействовал с ФДК, в нем запускается программа апоптоза. Предшественники плазмоцитов выходят из узелка через боковые зоны короны в интерфолликулярное плато, а затем перемещаются через паракортикальную зону в мозговое вещество (в мякотные тяжи), где созревают в плазмоциты.
В центре размножения находятся не только ФДК, но также CD4+ Т- лимфоциты (Т-хелперы/индукторы), которые взаимодействуют с ФДК. ФДК не экспрессируют МНС II класса, поэтому связывают иммунные комплексы (иккосомы) специальными рецепторами своей поверхности. Все эти клетки, а также специализированные макрофаги краевого синуса способствуют развитию В-клеточного иммунного ответа, в частности, иммунологической В- клеточной памяти - одной из главных функций центров размножения. Функциями центров размножения являются также: 2) клональная пролиферация В-лимфоцитов; 3) переключение класса иммуноглобулинов; 4) селекция лимфоцитов антигеном.
ПОСЛЕДОВАТЕЛЬНОСТЬ ИММУННЫХ РЕАКЦИЙ В В-ЗОНАХ ЛИМФО-
УЗЛА. Так как в лимфоидных узелках подвергаются превращениям в основном В-лимфоциты, то они являются В-зависимой зоной лимфоузла. При этом в центрах размножения вторичных лимфоидных узелков происходит активация В-лимфоцитов и созревание их аффинности (сродства) к антигену. Для этого необходимо, чтобы первичный узелок превратился во вторичный. Сигналом для превращения является поступление в первичный узелок либо 1) антигена, либо 2) антигенспецифических клеток (В-лимфоциты памяти). Первичные В-клетки памяти, экспрессирующие на своей поверхности иммуноглобулиновые рецепторы (sIg), поступают в лимфоидный узелок и покидают его либо в виде В-лимфоцитов памяти, либо в виде плазмоцитов. Эта их миграция может происходить с лимфой или с кровью через
368
посткапиллярные венулы с высоким эндотелием паракортикальной зоны как из вторичных узелков данного лимфоузла, так и из вторичных узелков других периферических органов иммуногенеза (пейеровы бляшки, аппендикс, миндалины, селезенка).
При развитии иммунного ответа по первому сценарию антигены поступают в лимфоузел с лимфой и распространяются по синусам. Они захватываются макрофагами и частично процессируются ими, а затем, поступая в В- зону, подвергаются дальнейшему процессированию фолликулярными дендритными клетками, которые выносят их на свою поверхность. ФДК осуществляют примирование Т-лимфоцитов СD4+ (Т-хелперов). Эти лимфоциты, кооперируя с В-лимфоцитами, передают им сигнал бласттрансформации, деления и дифференцировки в плазмоциты и В-лимфоциты памяти.
Во втором случае происходит следующее. Если первичный лимфоидный узелок заселяется хотя бы одной В-клеткой памяти, она превращается в первичный В-бласт и быстро формирует центр размножения с образованием вторичного узелка. В нем формируется темная зона, обращенная к паракортикальной зоне. Здесь В-бласты митотически делятся. В течение 4 суток их количество достигает примерно 10000. На 4-е сутки первичные бласты превращаются в центробласты, лишенные поверхностных иммуноглобулинов (sIg--клетки). Из центробластов образуются центроциты, которые перемещаются в светлую базальную зону. После взаимодействия центроцитов с фолликулярной дендритной клеткой происходит презентация антигена В-клеткам и активация последних. Они размножаются с образованием так называемых вторичных В-клеток-бластов. Клетки начинают экспрессировать на своей поверхности ген выживаемости (бессмертия) bcl-2 и смещаются в апикальную светлую зону центра размножения, а затем покидают его и превращаются либо в В-лимфоциты памяти, либо в плазмоциты. Эти клетки в последующем могут попасть в другие первичные узелки данного лимфоузла либо другого периферического органа иммуногенеза и запустить в них описанные выше процессы (такой способностью обладают прежде всего В-лимфоциты памяти). Это происходит и при вторичном иммунном ответе.
Если взаимодействие центроцитов с ФДК и их активация (селекция) антигеном не состоялись, клетки не экспрессируют ген бессмертия bcl-2, и в базальной светлой зоне центра размножения запускается программа их апоптоза. Здесь обнаруживаются макрофаги, осуществляющие фагоцитоз апоптозных тел.
ФДК имеют отростчатую форму и в области отростков связываются друг с другом десмосомами в густую сеть. Это исключает возможность их миграции из мест своей локализации. ФДК не экспрессируют на своей поверхности белков МНС II класса и поэтому фиксируют на своей поверхности частично процессированные макрофагами антигены с помощью рецепторов
369
к компонентам комплемента (CD21 и CD35). Эти рецепторы (иккосомы), связывая иммунные комплексы, сохраняют их на своей поверхности. ФДК экспрессируют также рецепторы к Fc-фрагменту иммуноглобулинов.
Таким образом, при гуморальном ответе информацию об антигене в В- зонах могут представлять макрофаги и ФДК.
Лимфоидные узелки лимфатических узлов являются весьма динамичными структурами. Они возникают при иммунном ответе и достаточно быстро исчезают при его затухании. В связи с этим различают несколько стадий развития узелков:
1.Формирование центра размножения;
2.Появление вокруг центра размножения короны из малых лимфо-
цитов;
3.Стадия максимального развития;
4.Стадия обратного развития.
На высоте иммунного ответа лимфоидные узелки достигают максимальной величины и могут смещаться в мозговое вещество.
ПАРАКОРТИКАЛЬНАЯ ЗОНА, ПКЗ (ЗОНА ГЛУБОКОЙ КОРЫ). Эта зона находится на границе коркового и мозгового вещества. Она является тимусзависимой зоной (Т-зоной) лимфоузла. ПКЗ содержит преимущественно Т-лимфоциты-хелперы, однако здесь обнаруживаются и мигрирующие в мякотные тяжи мозгового вещества плазмоциты на разных стадиях развития. После инъекции любого Т-зависимого антигена (антигена, требующего распознания как В-, так и Т-лимфоцитами) в паракортикальной зоне начинается активная пролиферация Т-лимфоцитов. Клетками микроокружения в паракортикальной зоне являются ретикулярные клетки фибробластического ряда, макрофаги и интердигитирующие клетки (ИДК).
ИДК имеют пальцевидные отростки, которые вдавливаются в многочисленные аналогичные клетки. В световом микроскопе при специальной окраске их с помощью реакции на фермент аденозинтрифосфатазу (АТФазу) видна сеть, сформированная этими клетками (см. рис. 21.3,Б). ИДК имеют светлые, неправильной формы ядра с краевым расположением хроматина. В цитоплазме клеток при электронной микроскопии обнаруживаются многочисленные везикулы, развитый комплекс Гольджи, гладкая ЭПС. Клетки образуют контакты со многими лимфоцитами. Интердигитирующие клетки являются трансформированными клетками Лангерганса эпидермиса кожи, которые транспортируют в лимфоузел антигены из кожи (очевидно, и из слизистой оболочки ротовой полости) в регионарный лимфоузел и в паракортикальной зоне трансформируются в ИДК. При этом они теряют гранулы Бирбека, свойственные клеткам Лангерганса. Подобная миграция клеток из кожи в регионарный лимфоузел является эффективным способом доставки антигена к Т-хелперам лимфоузлов. На ИДК обильно экспрессированы белки МНС II класса, необходимые для презентации антигена Т-
370