

2 курс / Гистология / Мяделец.Частная гистология
.pdfтакже структурные основы иммунных реакций, называется иммуноморфологией.
ИММУННАЯ СИСТЕМА. СОСТАВ И ФУНКЦИИ. Защитные реакции организма выполняются системой иммунитета. В ее состав входят:
1)Иммунокомпетентные органы;
2)Иммунокомпетентные ткани;
3)Иммунокомпетентные клетки, рассредоточенные по всему организму и выполняющие функции надзора за его антигенным гомеостазом. Иммунокомпетентные клетки продуцируют химические факторы иммунитета (антитела, цитокины и др.), участвующие в элиминации антигена.
Органы иммунной системы делятся на первичные и вторичные. К
первичным органам иммунитета относятся красный костный мозг, сумка Фабрициуса (у птиц) и тимус. Функцией первичных органов является осу-
ществление антигеннезависимой дифференцировки иммунокомпетентных клеток под воздействием специфических факторов - поэтинов, вырабатываемых стромой этих органов. При этом на поверхности иммунокомпетентных клеток происходит образование специфических для каждой популяции клеток рецепторов.
Вторичными органами иммунитета являются селезенка, лимфоузлы, миндалины, аппендикс, одиночные и групповые (пейеровы бляшки) лимфоидные узелки. В этих органах происходит антигензависимая дифференци-
ровка Т- и В-лимфоцитов, т.е. при непосредственном участии антигена
образуются иммунокомпетентные клетки, способные его уничтожать.
К иммунокомпетентным тканям относятся ретикулярная, лимфоидная, миелоидная ткани, а также кровь и в определенной степени рыхлая соединительная ткань. Сюда можно также отнести искусственно выделяемые ткани (на самом деле под ними понимается определенная совокупность иммунокомпетентных клеток): кожноассоциированную лимфоидную ткань (КАЛТ) и ассоциированную со слизистыми оболочками лимфоид-
ную ткань: бронхассоциированную лимфоидную ткань (БАЛТ), кишечноассоциированную лимфоидную ткань (КиАЛТ), лимфоидную ткань, ассоциированную с мочевой системой, лимфоидную ткань, ассоцииро-
ванную с конъюнктивой глаз и т.д. Эти “ткани” представлены лимфоидными узелками и диссоциированными иммунокомпетентными клетками. Они располагаются в области входных ворот инфекции и защищают организм от проникновения чужеродных агентов. Иммунокомпетентные ткани входят в состав органов иммунной системы.
Иммунокомпетентными клетками являются макрофаги, лимфоциты,
плазмоциты, все виды гранулоцитов, тканевые базофилы, натуральные киллеры. Их роль в иммунных реакциях организма будет рассмотрена ниже.
ФУНКЦИИ ИММУННОЙ СИСТЕМЫ. Иммунная система выполняет следующие функции:
331
1)Интегративная и регуляторная функции. Иммунная система отно-
сится к числу трех регуляторных систем организма наряду с нервной и эндокринной системами. С этими системами иммунная система тесно взаимодействует. Клетки и неклеточные (молекулярные) факторы иммунной системы оказывают регулирующее влияние на функции практически всех клеток, тканей и органов, участвуют в процессах эмбриогенеза и морфогенеза, дифференцировки и регенерации клеток и тканей. Обладая способностью к рециркуляции, клетки иммунной системы способны передавать информацию во все органы организма, участвуя в интеграции их в единый организм.
2)Функция сохранения и поддержания антигенного и тканевого го-
меостаза. Иммунная система элиминирует из организма все чужеродные агенты, как внедрившиеся извне, так и образовавшиеся в организме в результате трансформации. Она участвует также в регуляции деления и апоптоза клеток.
3)Барьерно-защитная функция - защита организма от возбудителей инфекционных (бактерии, их токсины, вирусы, грибки, паразиты) и неинфекционных (злокачественные опухоли, чужеродные трансплантаты и др.) агентов.
АНТИГЕНЫ. ПУТИ ПРОНИКНОВЕНИЯ АНТИГЕНОВ В ОРГАНИЗМ
Антигены - чужеродные вещества, способные при попадании в организм вызвать иммунные реакции. Различают полные и неполные анти-
гены. Полные антигены - белки, полисахариды, нуклеиновые кислоты и нуклеопротеины, гликопротеины, липопротеины, синтетические высокополимерные соединения, вирусы, паразиты, бактерии, чужие и собственные, но измененные действием внешних и внутренних факторов клетки и макромолекулы, тяжелые металлы. Полные антигены способны вызывать иммунный ответ без каких-либо дополнительных условий. Гаптены, или неполные антигены - низкомолекулярные соединения, которые сами по себе не могут вызвать иммунного ответа, однако соединяясь с клетками и тканями организма, изменяют их макромолекулы и в комплексе с такими измененными молекулами становятся полными антигенами.
Антигены могут проникать в организм разными путями: 1. Через дыхательные пути.
2. Через желудочно - кишечный тракт.
3. Через неповрежденную и особенно через поврежденную кожу.
4. Через слизистые оболочки глаз, носа.
5. Через слизистые оболочки мочеполовых путей.
6. Через кровь (при укусе насекомых, при инъекциях).
Пути проникновения антигена в организм называются входными воротами. В области входных ворот инфекции сосредоточены иммунокомпетентные клетки или ткани (например, лимфоидные узелки), а в непосредственной близости от них часто находятся вторичные иммунокомпетентные
332
органы (регионарные лимфатические узлы и др.). Кроме того, в этих участках действуют неиммунные факторы резистентности.
АНТИТЕЛА
Антитела - вещества белковой природы (иммуноглобулины, Ig), которые образуются в организме в ответ на антиген и, специфически взаимодействуя с ними, инактивируют их, обеспечивая гуморальный иммунитет. Антитела образуются активированными В-лимфоцитами и в особенности плазмоцитами - клетками, которые являются результатом конечной дифференцировки В-лимфоцитов. Антитела обладают специфичностью по отношению к антигену: они связываются только с тем антигеном, на который выработались. В образующемся при этом комплексе “антиген - антитело” антиген теряет свою биологическую активность. В дальнейшем этот комплекс подвергается разрушению клетками или неклеточными факторами иммунитета.
Существует несколько классов иммуноглобулинов: Ig G, М, А, Е, D. Наибольшее значение имеют иммуноглобулины класса G. Они составляют около 75% всех Ig сыворотки крови. Ig G имеют мономерное строение. Они нейтрализуют бактериальные токсины и вирусы. В иммунных реакциях Ig G вырабатываются при вторичном иммунном ответе и позже других антител. Они способны активировать систему комплемента по классическому пути, а также макрофаги и нейтрофилы. Ig G - единственный класс иммуноглобулинов, способный проникать через плацентарный барьер и участвовать в естественном пассивном иммунитете.
Иммуноглобулины класса М составляют около 10% всех Ig сыворотки крови. Они состоят из пяти связанных между собой молекул Ig, являясь пентамерами. IgM менее специфичны по отношению к антигенам, но более эффективны, чем IgG, при нейтрализации токсинов, активации комплемента, лизисе чужеродных клеток. IgM являются основными антителами, вырабатываемыми на ранних стадиях первичного иммунного ответа. Мономеры IgM являются одними из рецепторов В-лимфоцитов. Небольшая часть IgM может входить в связанном с секреторным компонентом виде в состав секретов экзокринных желез (секреторные IgM).
Иммуноглобулины А содержатся в биологических жидкостях: сыворотке крови, секретах экзокринных желез (слюне, слезе, слизи), моче и т.д.
Различают сывороточные и секреторные Ig A. Сывороточный IgA (10%
всех Ig крови) может существовать в димерной, тримерной и тетрамерной формах, причем преобладает димерная форма. Секреторный IgА вырабатывается плазматическими клетками лимфоидных узелков (фолликулов) и служит для защиты слизистых оболочек. В молекуле секреторного IgА два мономера связываются с так называемым секреторным компонентом, продуцируемым клетками эпителия. Секреторный компонент предохраняет IgA от действия агрессивных факторов секретов. IgА препятствует прикрепле-
333
нию микроорганизмов к поверхности эпителиев, а также играет роль опсонина: покрывая поверхность микроорганизмов, резко стимулирует фагоцитоз их фагоцитами.
Иммуноглобулины класса Е (реагины) участвуют в аллергических реакциях. Они вырабатываются в основном плазмоцитами слизистых оболочек воздухоносного и пищеварительного трактов. В крови присутствуют в крайне незначительных количествах (0,004%). Ig E вызывают выделение гистамина тучными клетками после того, как комплекс “антиген - антитело” присоединяется к их поверхности. Гистамин вызывает отек, сдавление сосуда, что уменьшает концентрацию антигена и препятствует его распространению.
Иммуноглобулины класса D составляют 1% от всех Ig. Их функции до конца не изучены. Известно, что они являются рецепторами В- лимфоцитов и играют важную роль в процессах их дифференцировки. Ig D могут фиксировать комплемент и часто направлены против антигенов, длительно циркулирующих в крови. Их количество может увеличиваться при некоторых вирусных инфекциях. Ig D выявляются в крови, секрете бронхов, молоке, спинномозговой жидкости.
Специфически взаимодействующие с антигенами иммуноглобулины любого из пяти классов называются антителами.
Для понимания механизма действия антител необходимо хотя бы вкратце рассмотреть их структуру (Рис. 21.1). Молекула антитела напоминает букву Y и состоит из двух тяжелых и двух легких цепей, связанных между собой дисульфидными мостиками. И тяжелые, и легкие цепи идентичны и расположены симметрично. Фермент папаин расщепляет молекулу антитела на два одинаковых Fab-фрагмента, каждый из которых имеет антигенсвя-
зывающий центр (fragment antigen binding, сокращенно Fab), и Fc-
фрагмент (fragment crystallizable), неспособный связывать антигены, но определяющий ряд биологических свойств антител: способность к связыванию комплемента, транспорту через плаценту, связыванию с иммунокомпетентными и неиммунными клетками, различными тканями. Структурное разнообразие антител определяется прежде всего последовательностью аминокислот в вариабельных областях тяжелых и легких цепей. Антитела участвуют в реакциях гуморального иммунитета - т.е. в реакциях, направленных против растворимых в жидкостях корпускулярных антигенов.
МЕХАНИЗМЫ ДЕЙСТВИЯ АНТИТЕЛ.
1. Простое связывание антигена Fab-фрагментами антитела выключает его действие на клетки, т.к. антитело закрывает активные, цитотоксические центры антигена. Образованный комплекс антиген-антитело в последующем выделяется с мочой, потом, слюной и т.д., т.е. элиминируется из организма.
334
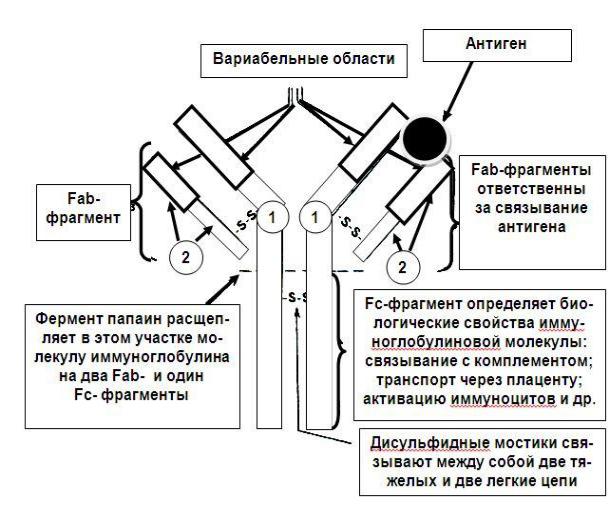
Рис. 21.1. Схема строения молекулы иммуноглобулина
2.Комплекс антиген - антитело при помощи Fс-фрагмента активирует макрофаги, моноциты и гранулоциты, которые включаются в элиминацию антигена путем фагоцитоза.
3.Покрывая поверхность чужеродной клетки, антитела класса G своими Fс-фрагментами активируют K-клетки (киллерные клетки). Эти клетки осуществляют антителозависимый лизис чужеродных клеток.
4.Комплекс антиген - антитело при помощи Fс-фрагмента активирует систему комплемента. Эта система включает ряд белков крови, которые составляют 10% от всех глобулинов крови. Данные белки, отличаясь от иммуноглобулинов, дополняют их в защитных реакциях. Активация системы комплемента имеет три важных следствия для защиты организма:
а) Белки комплемента, действуя по принципу перфоринов, встраиваются в цитолемму чужеродной клетки и образуют в ней сквозные каналы.
Врезультате клетка погибает от осмотического шока.
б) Компоненты комплемента активируют макрофаги, которые далее фагоцитируют антиген.
335
в) Белки комплемента вызывают местную воспалительную реакцию с активацией нейтрофильных лейкоцитов, уничтожающих антиген.
5.Иммуноглобулины класса Е осуществляют защитные реакции через систему тучных клеток (тканевых базофилов). В комплексе с антигеном они адсорбируются на поверхности этих клеток и вызывают их дегрануляцию. Содержащийся в гранулах гистамин вызывает расширение сосудов и отек, что уменьшает концентрацию антигена и препятствует его распространению
ворганизме. Одновременно гистамин вызывает сокращение гладких мышц полых органов, что ведет к механическому удалению антигена (например, паразитов) из организма. Кроме того, гепарин тучных клеток активирует макрофаги.
6.Антитела обладают собственной медленной ферментативной активностью (так называемая абзимная активность) и могут расщеплять некоторые субстраты, в том числе и антигенного характера.
ИММУНОКОМПЕТЕНТНЫЕ КЛЕТКИ
К иммунокомпетентным относятся Т- и В - лимфоциты, плазмоциты,
макрофаги, тучные клетки (тканевые базофилы), гранулоциты, NКклетки. По функциональному признаку все эти клетки делятся на не-
сколько групп:
1. Антигенпредставляющие клетки.
2.Эффекторные клетки.
3.Регуляторные клетки.
4.Вспомогательные клетки.
5.Клетки иммунологической памяти.
АНТИГЕНПРЕДСТАВЛЯЮЩИЕ КЛЕТКИ (АПК). Это клетки, которые
передают информацию об антигене Т- и В-лимфоцитам. Они рассредоточены на главных путях поступления антигена в организм: в эпидермисе и дерме кожи, в эпителии и собственной пластинке слизистых оболочек, а также в периферических органах иммунной системы. К антигенпредставляющим клеткам относятся:
1.Макрофаги и моноциты.
2.Интердигитирующие клетки (ИДК).
3.Фолликулярные дендритные клетки (ФДК) лимфоидных узелков
4.В-лимфоциты.
5.М-клетки эпителия пейеровых бляшек.
В последнее время специализированными АПК считаются дендритные клетки (см. ниже).
Представление (презентация) антигена протекает в несколько этапов
(Рис. 21.2):
1. Фагоцитоз (пиноцитоз) неизмененных антигенов;
336
2.Процессинг (переработка) антигена. Он заключается в частичном протеолизе антигенных молекул при помощи ферментов лизосом в гуморальном или протеасом – в клеточном иммунитете (см. ниже). При этом высвобождаются и концентрируются высокоиммунные антигенные детерминанты, представляющие собой линейные пептидные цепочки длиной в 8-11 аминокислот. Остальные части молекулы антигена расщепляются лизосомальными ферментами до простых соединений. Процессинг антигена протекает в кислой среде и длится около 1 часа.
3.Биосинтез молекул главного комплекса гистосовместимости
(Major Histocompatibiliti Complex, или MHC). Эти молекулы у человека часто называются HLA-антигенами (от англ. Human Leukocyte Аntigens -
человеческие лейкоцитарные антигены). Различают два класса молекул МНС: I и II класса.
Молекулы МНС I класса обнаруживаются во всех клетках организма
иявляются их специфическим антигенным “паспортом”. Эти антигены воспринимаются как чужеродные иммунокомпетентными клетками другого индивидуума. Они синтезируются в гранулярной ЭПС. Кроме того, в качестве антигенов при клеточных иммунных реакциях выступают эндогенные антигены - белковые молекулы вирусов, внутриклеточных паразитов, раковых клеток и др. Эти антигены подвергаются процессингу (АТФ-зависимому протеолизу) в специальных комплексах ферментов-протеаз – протеосомах. Образующиеся полипептидные цепочки с помощью специальных белков-
переносчиков (ТАР- белки - от Transporter for Antigen Presentation) перено-
сятся в гранулярную ЭПС, соединяются с молекулами МНС I класса, поступают в комплекс Гольджи, упаковываются в мембранные пузырьки и далее экспрессируются на поверхности плазмолеммы АПК. Их распознают цитотоксические Т-лимфоциты. Благодаря этим молекулам цитотоксические Т- лимфоциты узнают, а затем уничтожают все клетки, приобретшие или имеющие свойство чужеродности для организма: инфицированные, опухолевые клетки, а также клетки трансплантатов.
Молекулы МНС II класса синтезируются и экспрессируются только АПК. Они синтезируются в гранулярной эндоплазматической сети, далее поступают в комплекс Гольджи, заключаются в мембранные пузырьки. Экзогенные антигены захватываются АПК с помощью эндоцитоза. Далее в эндосоме происходит процессинг антигена. Затем эндосома сливается с мембранным пузырьком, происходит связывание фрагментов антигена с МНС II,
иэтот комплекс экспрессируется на плазмолемме.
4. Выделение процессированного антигена на поверхность АПК и презентация его лимфоцитам (см. выше). Соединение молекул МНС I или II класса с процессированным и переведенным в высокоиммунную форму антигеном и выделение образованного высокоактивного иммунного комплекса на
337
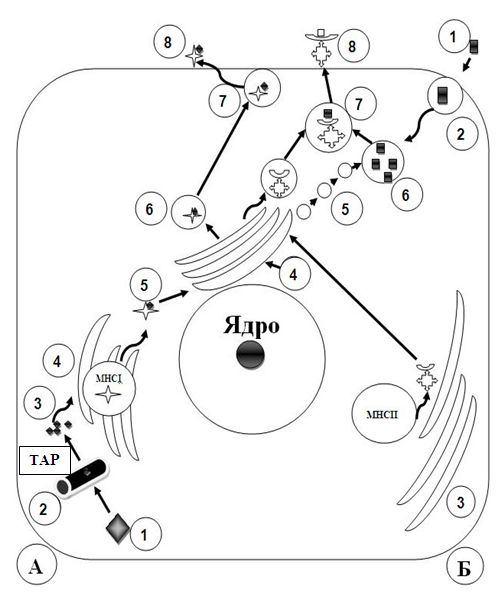
Рис. 21.2. Презентация антигенов при иммунном ответе А – клеточный иммунитет: 1 – антиген в генетически чужеродной клетке (напри-
мер, инфицированной вирусом) процессируется в протеосоме 2 и в процессированной форме 3 с помощью ТАР-белков поступает в гранулярную ЭПС 4, где синтезируются молекулы МНС-I. После связывания антигена с МНС-I (5) комплекс АГ+ МНС-I поступает в комплекс Гольджи, где упаковывается в мембранные пузырьки 6, которые транспортируют этот комплекс и выделяют на поверхность клетки (7,8) Б – гуморальный иммунитет: 1 – антиген; 2 – эндоцитоз антигена; 3 – биосинтез в гранулярной ЭПС молекул MHC-II и транспорт их в комплекс Гольджи 4. Здесь молекулы МНС-II упаковываются в мембранные пузырьки; 5 - лизосомы, образованные в комплексе Гольджи 4, направляющиеся к эндосоме с антигеном и сливающиеся с ней с образованием эндолизосомы. Происходит процесссинг антигена (6); 7 – конъюгация процессированного антигена с МНС-II осуществляется после слияния эндолизосомы с транспортным пузырьком для МНС-II; 8 – экспрессия комплекса АГ+ МНС-II на плазмолемме АПК
338
поверхность АПК дает возможность включаться в иммунные реакции лимфоцитам – основным иммунокомпетентным клеткам.
5. Выработка и секреция АПК медиаторов, стимулирующих Т-лимфоциты- хелперы и усиливающих их пролиферацию. Наиболее важными среди них являются интерлейкин -1 (ИЛ-1), ИЛ-12, ИЛ-18, фактор некроза опухоли
α (ФНОα).
Если антиген в ходе фагоцитоза макрофагами полностью разрушается и удаляется из организма, то иммунная реакция на этом заканчивается. Однако гораздо чаще имеют место ситуации, когда макрофаги не могут полностью самостоятельно ликвидировать антиген. В таком случае антигенпредставляющие клетки привлекают к уничтожению антигена лимфоциты.
Морфофункциональная характеристика различных АПК
МАКРОФАГИ. Макрофаги образуются из моноцитов крови путем специфической дифференцировки. Эта дифференцировка происходит в тканях и сопровождается увеличением размеров клетки до 25-50 мкм, приобретением ядром бобовидной формы, накоплением лизосом и других органелл: митохондрий, эндоплазматической сети. Гипертрофируется комплекс Гольджи, нарастает количество пиноцитозных пузырьков. Резко усложняется поверхность макрофага за счет увеличения количества микроворсинок и складок, возрастают подвижность макрофагов, способность их к пиноцитозу и фагоцитозу, бактерицидная активность. В клетках нарастает активность лизосомальных ферментов, а также ферментов энергетического обмена. Ферменты лизосом осуществляют обработку антигена и его разрушение до мелких высокоиммунных комплексов. Эти комплексы связываются с молекулами МНС II класса и выходят на поверхность макрофага. Далее с ними контактируют Т- и В-лимфоциты. Наиболее активны в отношении презентации антигена лимфоцитов макрофаги краевого синуса лимфоузлов, селезенки, печени.
Обработка антигена и передача его лимфоцитам - наиболее важная функция макрофагов. Кроме того, они выполняют ряд других важных функций.
Секреторная функция. Макрофаги секретируют: бактерицидные вещества; ряд ферментов, оказывающих бактерицидное действие; интерфероны, участвующие в противовирусном иммунитете; компоненты компле-
мента; факторы, стимулирующие размножение лимфоцитов (ИЛ-1), кле-
ток - предшественниц миелопоэза, фибробластов; тромбоцитарный фактор роста. Вместе с тем, макрофаги выделяют и вещества, подавляющие деление лимфоцитов, опухолевых клеток (фактор некроза опухолей α) и стимулирующих их апоптоз. Из других синтезируемых макрофагами следует назвать
простагландины и лейкотриены, компоненты комплемента, свободные радикалы кислорода, перекись водорода, оксид азота, катионные белки.
Эти вещества либо обладают бактерицидным и цитоцидным действием, либо участвуют в воспалительных реакциях.
339
Функция фагоцитоза. Макрофаг может фагоцитировать и разрушать антиген. Он фагоцитирует и крупные частицы, не являющиеся антигенами, например, частицы угля, пыли и др. Накопление в цитоплазме макрофага этих инертных веществ подавляет его фагоцитарную активность и другие иммунные функции.
На своей поверхности макрофаги несут специфические рецепторы: к Fсфрагменту антител; к С3-компоненту комплемента; к антигенам и гормонам. Для макрофагов характерен специфический репертуар CD-антигенов. Среди них наибольшее значение имеет CD14. СD14-молекулы связывают липополисахариды бактерий.
ДЕНДРИТНЫЕ КЛЕТКИ ЛАНГЕРГАНСА (ДКЛ) ЭПИДЕРМИСА И МНОГОСЛОЙНЫХ ЭПИТЕЛИЕВ (Рис. 21.3, А). В эпидермисе количество КЛ составляет примерно 3-10% от всех кератиноцитов (эпителиоцитов). У человека КЛ локализуются преимущественно в шиповатом, иногда в базальном слоях эпителиев. Встречаются также в дерме. Кроме многослойных эпителиев, КЛ обнаруживаются в многорядном эпителии воздухоносных путей. Они имеют костномозговое происхождение, развиваясь из CD34+ -клетки костного мозга. В цитоплазме КЛ имеют развитые органеллы белкового синтеза, лизосомы. а также специфические гранулы – фляжкообразные полосатые пузырьки, имеющие форму теннисной ракетки. Предполагают, что эти гранулы участвуют в транспорте антигена. КЛ могут мигрировать в регионарные лимфоузлы, превращаясь в интердигитирующие клетки. На поверхности КЛ несут белки МНС I и II класса. Клетки играют роль антигенпредставляющих клеток в многослойных эпителиях, дерме и собственной пластинке слизистых оболочек. Они осуществляют также транспорт антигена на своей поверхности в регионарные лимфоузлы.
ИНТЕРДИГИТИРУЮЩИЕ КЛЕТКИ (ИДК). ИДК (Рис. 21.3, Б). являются разновидностью макрофагов. Они располагаются в Т-зависимых зонах периферических органов иммунитета: паракортикальных зонах лимфоуз-
лов, в периартериальных зонах селезенки, интерфолликулярных зонах лимфоидных скоплений (пейеровых бляшек, миндалин, аппендикса).
Считается, что ИДК образуются из клеток Лангерганса эпидермиса и других, мигрирующих из кожи и слизистых оболочек в лимфоузел. В процессе миграции происходит изменение фенотипических черт клеток. ИДК имеют длинные отростки, которые контактируют с большим числом Т-лимфоцитов. Они способны перерабатывать антиген, выделять его на свою поверхность, а затем представлять Т-лимфоцитам. Антиген на поверхности ИДК может сохраняться долгое время, и в результате контакта с ним постоянно вырабатывается какое-то количество эффекторных Т-лимфоцитов, способных уничтожать антиген. Поэтому при повторном попадании антигена в организм иммунная реакция идет значительно быстрее. Таким образом, ИДК участ-
вуют в иммунной памяти.
340