

2 курс / Гистология / КОЛИЧЕСТВЕННАЯ_ОЦЕНКА_ИЗМЕНЕНИЙ_В_МИКРОСТРУКТУРЕ_ПЕРИНЕЙРОНАЛЬНЫХ
.pdfисследовательские стипендии – Краткосрочные стипендии»; средствами РФФИ в рамках конкурса «Аспиранты» (проект 20-315-90074); стипендией Правительства Российской Федерации для обучающихся по образовательным программам высшего образования по очной форме по специальностям или направлениям подготовки, сутствсоответствующим приоритетным направлениям модернизации и технологического развития российской экономики.
Публикации.
По материалам диссертации опубликовано 15 работ, из них 5 – статьи в рецензируемых изданиях и представленных в базах данных Scopus и Web of
Science, 10 – тезисы докладов.
Структура и объем работы
Диссертация состоит из введения, 3 глав, заключения и выводов,
списка публикаций автора, списка цитируемой литературы из 188
наименований. Работа изложена на 129 страницах, содержит 49 рисунков.
Во введении обоснована актуальность диссертационной работы,
сформулированы цель и задачи, приведены объекты и методы их исследования, отмечены научная новизна и практическая ценность полученных результатов, положения, выносимые на защиту.
В первой главе представлен обзор литературы. Описаны история исследования, состав, пространственная структура и функции PNN, а также механизмы влияния PNN на регенерацию и пластичность ЦНС.
Во второй главе дан обзор литературы по методам исследования PNN,
использованные при выполнении работы, в том числе эпифлуоресцентная микроскопия низкого разрешения и конфокальная микроскопия высокого разрешения, а также разработка методов анализа данных микроскопии.
В третьей главе представлены разработанные нами новые методы и полученные на их основе экспериментальные результаты.
11
https://t.me/medicina_free
Благодарности
Автор выражает благодарность научным руководителям – профессору КПФУ А.В. Аганову и доценту Университета г. Хельсинки М.Н. Павельеву, а
также соавторам исследования – профессорам А.П. Киясову, Х. Раувала,
Ю.Н. Осину, научным сотрудникам, студентам и лаборантам: Н. Арнсту, А.
Мельниковой, А. Двоеглазовой, А. Кочневой, Н. Шайхутдинову, А.
Жигалову, Н. Кулесской, Х. Яалиноя, С. Федосимовой, М. Мавликееву, П.
Уварову, Т. Балтиной, А. Титовой, Р. Каушику. Автор также выражает благодарность профессору А.Э. Дитятеву из Германского Центра Изучения Нейрозаболеваний за сотрудничество.
12
https://t.me/medicina_free
Глава 1. ПРОСТРАНСТВЕННАЯ СТРУКТУРА И
МОЛЕКУЛЯРНЫЙ СОСТАВ ПЕРИНЕЙРОНАЛЬНЫХ СЕТЕЙ
1.1. История исследования PNN
Первоначальные исследования PNN связаны с именем итальянского нейробиолога Камилло Гольджи, который в 1898 году описал как эндоклеточный ретикулярный аппарат (аппарат Гольджи), так и периферический ретикулум, расположенный на поверхности нервных клеток.
По мнению Гольджи эта оболочка являлась корсетом из «нейрокератина»,
задача которого останавливать «нервные потоки» между клетками [16]. С.
Рамон-и-Кахаль считал, что Гольджи необходимо поблагодарить за привлечение внимания к перинейрональным сетям и предоставление нам первого точного описания этих объектов [17].
Работы учеников Гольджи Э. Вератти [18] и А. Донаджио [19, 20, 21, 22, 23] показали, что перинейрональные сети состоят из многоугольных единиц. Предполагалось, что через эти пространства проходят терминали аксонов и соединяются с поверхностью нервных клеток [20, 24].
Следующий этап исследований PNN был связан с выделением двух типов внеклеточного матрикса ЦНС: традиционная перинейрональная сеть
(сеть Гольджи) и диффузная сеть (или «Füllnetz»), охватывающая всю нервную систему (Бете [25]). И перинейрональные, и диффузные сети являются частью синтициальной организации нейроглии согласно представлениям Г. Хельда [24]. В дальнейшем эта теория разрабатывалась рядом исследователей: Альцгеймер [26], Адамкевич [27], Беста [28],
Белчовски [29]. Значительный интерес вызывали также структурно-
функциональные связи между этими двумя типами сетей внеклеточного матрикса, глией и нейронами (К. фон Экономо [30]). С. Рамон-и-Кахаль [31,
17], однако, не соглашался с мнением Бете и утверждал, что перинейрональная сеть представляет собой экспериментальный артефакт,
возникающий в процессе коагуляции околоклеточной жидкости.
13
https://t.me/medicina_free
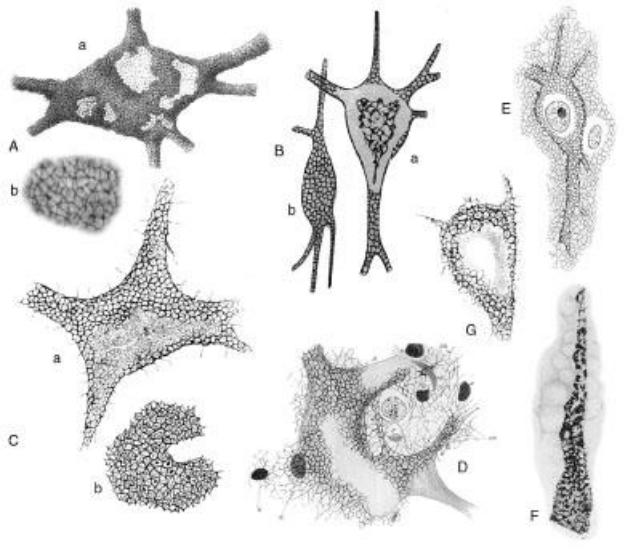
Рисунок 1. Иллюстрации перинейрональной сети, созданные разными авторами в течение столетия. Заимствовано из Celio et. al., «Perineuronal nets. Past and present» (1998) [32]. (А)
Нервные клетки с ретикулярной оболочкой (передний рог спинного мозга): (а) и (b),
увеличенные детали (а) иллюстрируют тонкую текстуру перинейрональной сети. (B) Две церебральные клетки с короткими аксонами (взрослая кошка), окрашенные восстановленным нитратом серебра, после фиксации в формоацетоне: (a) клетка с восходящим аксоном, рассматриваемая в экваториальной плоскости, (b) клетка с нисходящим аксоном, видимая в поверхностной плоскости. (C) Нервная клетка,
полученная из передних рогов спинного мозга собаки (a); Структура колесика (raggere di Donaggio), образованная тонкими нитями, исходящими из центрального пятна,
распознаваемого в сетках перинейрональной сети (b). Эти структуры, вероятно,
представляют собой усохшие синаптические окончания, которые занимают ячейки сети.
(D) Клетка с сетью Гольджи и диффузной сетью (передний рог спинного мозга эмбриона теленка), окрашенные по методу Бете (метиленовый синий Эрлиха и молибдат аммония).
14
https://t.me/medicina_free
(E) Кортикальная клетка взрослой собаки, окрашенная по модифицированному методу Бете. (F) Изменения в перинейрональной сети кортикальной клетки человека, полученные от пациента с паралитической деменцией. (G) Клетка, полученная из ядра блуждающего нерва (продолговатого мозга) Lacertamuralis. Внутри ячеек периферического аппарата
(или перинейрональной сети) виден типичный рисунок, исходящий из небольшого центрального пятна. Филаменты, исходящие с поверхности (клетки), соединяются с окружающей стромой.
В течение нескольких последующих десятилетий внимание микробиологов сместилось с ретикуляционных теорий к нейронной доктрине. Но с начала 30-х годов XX в. продолжались сравнительные исследования перинейрональных сетей в ЦНС земноводных рептилий и птиц
(Р. Рондинини [33]). Г. Беллони [34] показал, какие структурные изменения возникают в коре мозга человека при дегенеративных церебропатиях.
Благодаря этим исследованиям многие ученые перестали рассматривать PNN
как артефакт несовершенных морфологических методов.
В 1950-60-е гг. XX в. с развитием гистохимических методов исследования был обнаружен периодический кислотно-шифф-
положительный (PAS-позитивный) материал вокруг нервных клеток и высказано предположение о его связи с сетями Гольджи [35, 36, 37, 38].
Однако оно не находило подтверждения. Длительное время исследовалась связь этого материала с гематоэнцефалическим барьером [39], что привело к продолжительному отрицанию существования внутритканевого вещества в ЦНС. Однако в настоящее время существование внушительной внутритканевой полости в головном мозге, пронизывающей всё пространство
[40, 41] и заполненного молекулами внеклеточного матрикса [42, 43, 44], не вызывает сомнения.
1.2. Состав и пространственная структура PNN
Существуют два типа внеклеточного матрикса ЦНС: диффузный,
который находится во всей ЦНС, и конденсированный матрикс, PNN,
15
https://t.me/medicina_free
который находится вокруг нейронов. Оба типа содержат несколько типов хондроитинсульфат протеогликанов (ХСПГ) и длинные цепи гиалуронана, а
также тенасцин-С и тенасцин-R. PNN помимо этих молекул содержит один или нескольких связывающих белков (Crtl1 / Hapln1 и Bral2 / Hapln4) и
большие количества тенасцина-R. Этот ансамбль образует плотную структуру [45, 46, 47, 48, 49, 50]. Некоторые молекулы, обнаруживаемые в
PNN (гиалуронан-синтаза, аггрекан, нейрокан, связывающие белки),
вырабатываются нейронами, а другие - окружающей глией [45, 46].
ХСПГ являются основным компонентом PNN, в ЦНС они могут взаимодействовать с различными факторами роста и молекулами клеточной адгезии, играя значительную роль в развитии [51, 52]. В основном они оказывают ингибирующее действие на разрастание и регенерацию нейритов
[53]. Экспрессия ХСПГ активируется после повреждения ЦНС [54, 55, 56, 57, 58], а ферментативное расщепление ГАГ-цепей ХСПГ при помощи хондроитиназы ABC улучшает регенерацию аксонов и восстановление их функций [59, 60]. Например, установлено, что деградация ХСПГ вызывает рост аксонов Пуркинье в мозжечке [61] и способствует прорастанию волокон сетчатки после денервации верхнего колликула у взрослых крыс [62].
Расщепление ХСПГ восстанавливает пластичность после критического периода в зрительной коре взрослого человека [2].
У взрослых животных инъекция хондроитиназы АВС в зрительную кору восстанавливала пластичность глазного доминирования и позволила исправить амблиопию [2]. В спинном мозге введение хондроитиназы АВС способствовало функциональному восстановлению после повреждения и после неточного повторного соединения периферических аксонов [50; 63].
Также она способствовала регенерации аксонов и прорастанию коллатеральных клеток в конусообразном ядре и спинном мозге [59, 64]. Эти и другие эксперименты легли в основу предположения, что PNN играют центральную роль в контроле пластичности в ЦНС и в ограничении способности взрослой ЦНС восстанавливаться после повреждения.
16
https://t.me/medicina_free
Разновидности ХСПГ, которые отвечают за контроль пластичности,
пока не определены, но исследования животных, лишенных бревикана и нейрокана, выявили аномалии в одной форме пластичности - долгосрочной потенциации [11].
Основой структуры внеклеточного матрикса является гиалуроновая кислота (или гиалуронан) (HA), который присутствует во всех PNN, и все нейроны с PNN экспрессируют фермент гиалуронан-синтазу (HAS). В
исследованиях [45, 50] было показано, что ни один из известных HA-
связывающих белков не был обнаружен на поверхностях нейронов во время формирования PNN. В связи с этим авторы высказали предположение, что трансмембранный HAS или неопознанный рецептор HAS, присутствующий на поверхности PNN-несущих нейронов, действует как док, который иммобилизует PNN на поверхности нейронов.
Гиалуроновая кислота является линейным полимером из дисахаридных звеньев N-ацетилглюкозамина и D-глюкуроновой кислоты, соединенных β- 1,4- и β-1,3-гликозидными связями. Образование крупных агрегатов во внеклеточном матриксе обусловлено свойством гиалуроновой кислоты связываться с другими молекулами. При помощи гиалуронан-синтаз цепи гиалуроновой кислоты могут прикрепляться к плазматической мембране нейронов [45]. Предположительно, длина и скорость синтеза цепей гиалуроновой кислоты влияют на некоторые свойства PNN. Однако в настоящее время механизмы этого влияния не установлены.
ХСПГ в PNN принадлежат семейству лектиканов (аггрекан, нейрокан,
версикан и бревикан) и имеют глобулярный домен связывания HA,
предположительно связывающийся с перицеллюлярным гиалуронансином,
синтезированным трансмембранным HAS. В исследовании [65] было высказано предположение, что эти звенья ХСПГ/HA, как в хряще,
стабилизируются связывающими белками. Карбоксильные группы этих ХСПГ затем связываются с тенасцинами, которые, в свою очередь,
связываются с гликозаминогликановыми цепями хондроитинсульфатов,
17
https://t.me/medicina_free
вместе образуя массивную макромолекулу. Наиболее распространенным лектиканом является аггрекан, присутствующий почти во всех PNN, другие члены семейства встречаются только в определенных областях нервной системы [50]. Структурная целостность PNN усиливается за счет способности тримера тинасцина-R связывать до трех молекул лектикана, в
первую очередь аггрекана.
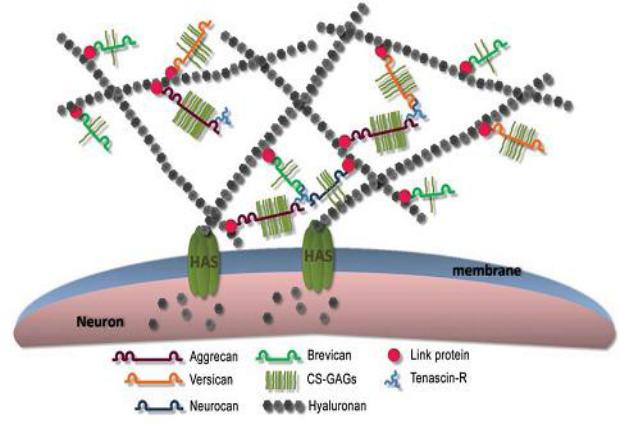
Рисунок 2. |
Структура перинейрональной сети. Заимствовано из Kwok J. C. F. |
|
«Extracellular matrix and perineuronal nets in CNS repair» (2011) [8] |
||
Чтобы |
понять, как формируются PNN, в |
исследовании, |
представленном в работе [50], была сделана попытка найти корреляцию между временем экспрессии различных молекул PNN относительно стадии развития PNN. Оно показало, что большинство компонентов экспрессируются задолго до того, как начинают формироваться PNN, но связывающий белок и аггрекан активируются одновременно с началом образования PNN. Животные, у которых в ЦНС отсутствует белок Crtl1,
имеют только рудиментальные PNN [66].
18
https://t.me/medicina_free
Бревикан представляет собой протеогликан, специфичный для головного мозга. Он увеличивает инвазивность клеток глиомы in vivo, и
предполагается, что играет определенную роль в развитии волокон центральной нервной системы. Чтобы изучить роль бревикана в развитии и функционировании мозга, авторы статьи [11] создали мышей, у которых отсутствовал функциональный ген бревикана. Эти мыши были жизнеспособны и плодовиты и имели нормальную продолжительность жизни. Анатомия мозга была нормальной, хотя были обнаружены изменения в экспрессии нейрокана. PNN формировались, но оказались менее заметными у мутантных мышей, чем у мышей дикого типа (контрольных). У мышей с дефицитом бревикана наблюдался значительный дефицит в поддержании долговременной потенциации гиппокампа (LTP). Однако явных нарушений возбуждающей и тормозной синаптической передачи обнаружено не было,
что указывает на сложную причину дефекта LTP. Детальный поведенческий анализ не выявил статистически значимых нарушений обучения и памяти.
Эти результаты демонстрируют, что бревикан не имеет решающего значения для развития мозга, но играет ограниченную структурную и функциональную роль.
В настоящее время микроструктура PNN остается практически неисследованной. Между тем, можно предположить, что пространственное распределение ХСПГ в PNN контролирует некоторые функции синапсов. Ряд данных указывает на то, что структуры из ХСПГ на поверхности нейронов нужны для упорядоченного распределения протеогликан-связывающих лигандов Sema3A, Otx2 и тенасцинов [45, 67, 68, 69].
Возможно, структура PNN контролирует распределение нейротрофических факторов BDNF, NT-3, FGF, VEGF, плейотрофина и лигандов семейства GDNF, связанных с высокой аффинностью с гликозаминогликанами, на поверхности нейронов [70, 71, 72].
В статье [15] авторы использовали подход, комбинирующий структурированную иллюминирующую микроскопию со сверхразрешением
19
https://t.me/medicina_free
(SR-SIM) и математическое реконструирование, чтобы количественно измерить топологию PNN. Было предположено, что восстановление после инсульта потребует обратимого разрыхления PNN, поскольку перинейроальный матрикс способен ограничивать нейронную пластичность,
но в то же время необходим для поддержания жизни синапсов. Результаты показали, что очаговая церебральная ишемия вызывает частичное истощение
PNN, и что умеренная гипоперфузия, не связанная с ишемической травмой,
может вызвать ультраструктурные перестройки в неповреждённых сетях. Это предполагает, что топологическое преобразование PNN может поддерживать перестройку нейронных сетей после инсульта.
1.3. Функции PNN
По всей видимости, из-за их большого вклада в нейронную пластичность [1, 2], функции PNN подвергаются изменению при многих неврологических заболеваниях, включая эпилепсию [73, 74], шизофрению
[75], травматическое повреждение мозга [76, 77] и инсульт [78].
Окончание критического периода пластичности связывается с завершением формирования PNN. Разрушение PNN в зрелом возрасте приводит к ювенильно-подобному состоянию пластичности, в котором возможно прорастание аксона и восстановление функции поврежденных нейронов (например, после травмы спинного мозга) [59, 64]. При генетическом удалении компонентов PNN в мозге грызунов в зрелом возрасте сохраняется синаптическая пластичность и появляется аномальная синаптическая передача [11].
PNN выполняют функцию буфера, поглощая катионы, что стабилизирует среду вокруг нейронов с высокой электрической активностью и значительными колебаниями уровня ионов. Поглощение достигается за счет большого некомпенсированного отрицательного заряда, обусловленного наличием сульфатных групп ГАГ-цeпей в составе PNN [5, 6, 79, 80, 81, 82].
20
https://t.me/medicina_free