

Ацетил-КоА карбоксилаза
АТФ АДФ + Фн
Ацетил-КоА |
|
биотин |
Малонил-КоА |
|
СО2 |
||||
|
|
|||
|
+ Цитрат, инсулин |
|
||
|
- |
Адреналин, глюкагон, |
|
|
|
- |
пальмитоил-КоА |
|
|
2стадии:
1.СО2 + биотин + АТФ → биотин-СООН + АДФ + Фн
2.ацетил-КоА + биотин-СООН → малонил-КоА + биотин
Регуляторная реакция синтеза пальмитата
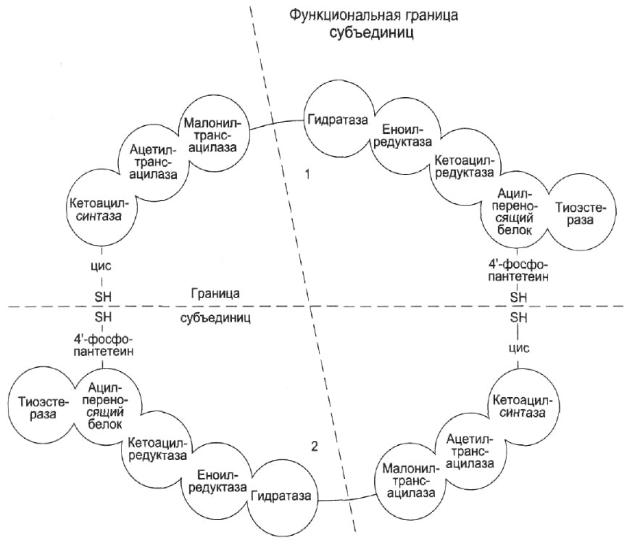
Пальмитатсинтетазный комплекс
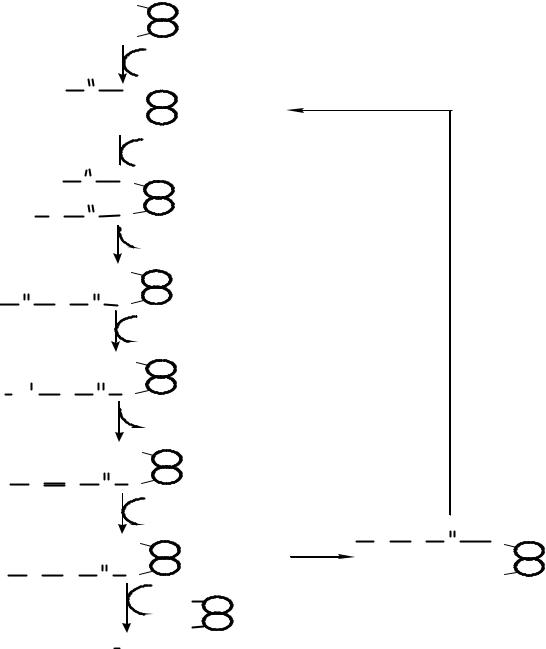
HS |
ÀÏ Á |
ï àëüì è ò àò ñè í ò åò àçí û é |
|
êî ì ï ëåêñ |
|
HS |
Ацети л-Ко А |
|
|
||
|
àöåò è ëò ðàí ñàöè ëàçí û é öåí ò ð |
|
O ÍSKoA
ÍSKoA
(R)H3C C S 
Àöè ë-ÀÏ Á
HS 
Ì àëî í è ë-Êî À
ì àëî í è ëò ðàí ñàöè ëàçí û é öåí ò ð
O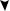
 ÍSKoA
ÍSKoA
(R)H3C |
C |
S |
Àöè ëì àëî í è ë-ÀÏ Á |
|
|
|||
|
|
H2 |
O |
|
|
|
||
HOOC |
C |
S |
|
|
|
|
||
C |
êåò î àöè ëñè í ò àçí û é öåí ò ð |
|
|
|||||
|
|
|
|
|
|
|
||
|
|
|
|
|
ÑÎ 2 |
|
|
|
|
|
|
HS |
|
|
|
|
|
|
Î |
H |
O |
|
Êåòî àöè ë-ÀÏ Á |
|
|
|
(R)H3C |
C |
C2 |
C |
S |
Í ÀÄÔÍ |
|
|
|
|
|
|
|
|
2 |
|
|
|
|
|
|
|
кет о аци лредукт азн ы й цен т р |
|
|
||
|
|
|
|
|
Í ÀÄÔ+ |
|
|
|
|
|
|
HS |
|
|
|
|
|
|
ÎH |
H |
O |
|
Î êñè àöè ë-ÀÏ Á |
|
|
|
(R)H3C CH |
C2 |
C |
S |
|
|
|
|
|
|
|
|
|
ãè äðàò àçí û é öåí ò ð |
|
|
||
|
|
|
|
|
Í 2Î |
|
|
|
|
|
|
HS |
|
|
|
|
|
|
H |
H |
O |
|
Åí î è ë-ÀÏ Á |
|
|
|
(R)H3C |
C |
S |
|
|
|
|
||
C |
C |
Í ÀÄÔÍ |
|
|
|
|||
|
|
|
|
|
2 |
|
|
|
|
|
|
|
ен о и лредукт азн ы й цен т р |
|
|
||
|
|
|
|
|
Í ÀÄÔ+ |
H |
H |
O |
|
|
|
HS |
|
(R)H3C C2 |
C2 |
C S |
|
(R)H3C |
H |
H |
O |
|
Àöè ë-ÀÏ Á |
|
Àöè ë-ÀÏ Á |
|
C2 |
C2 C |
S |
Í 2Î |
|
|
HS |
||
т и о эст еразн ы й цен т р |
|
|
|
|
||||
|
HS |
ÀÏ Á |
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
HS |
|
|
|
|
|
|
R CÎÎÍ |
|
|
|
||
ï àëüì è òàò
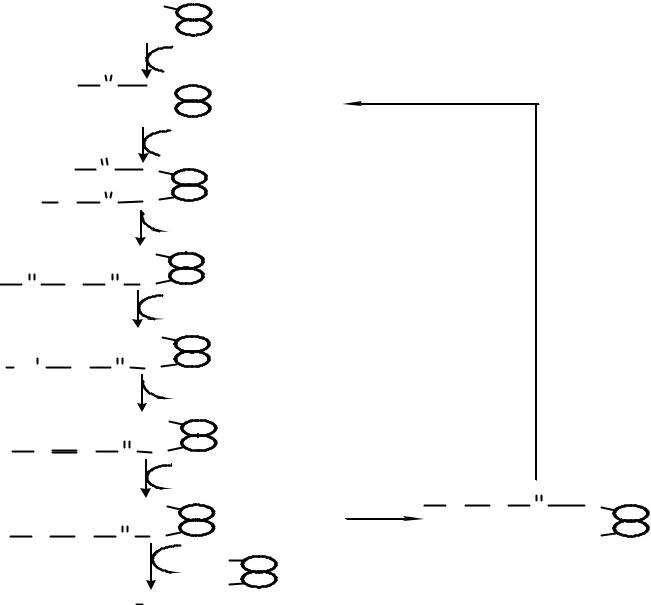
HS |
ï àë üì è ò àò ñè í ò åò àçí û é |
ÀÏ Á |
êî ì ï ë åêñ |
HS 
Ацети л-Ко А
àöåò è ë ò ðàí ñàöè ë àçí û é öåí ò ð
O ÍSKoA
ÍSKoA
(R)H 3C C S 
Àöè ë -ÀÏ Á
HS 
Ì àëî í è ë-Êî À
ì àë î í è ë ò ðàí ñàöè ë àçí û é öåí ò ð
O ÍSKoA
ÍSKoA
(R)H 3C |
C |
S |
Àöè ëì àëî í è ë-ÀÏ Á |
|
|
H2 |
O |
|
|
HOOC |
C |
S |
|
|
C |
êåò î àöè ë ñè í ò àçí û é öåí ò ð |
|||
|
|
|
|
|
|
|
|
|
ÑÎ 2 |
|
|
HS |
|
|
Î |
H2 |
O |
|
Êåòî àöè ë -ÀÏ Á |
(R)H3C C |
C |
S |
|
|
C |
Í ÀÄÔÍ 2 |
|||
|
|
|
|
|
|
|
|
|
кет о аци л редукт азн ы й цен т р |
|
|
|
|
Í ÀÄÔ + |
|
|
|
HS |
|
ÎH |
H2 |
O |
|
Î êñè àöè ë -ÀÏ Á |
(R)H 3C CH |
C |
S |
|
|
C |
|
|||
ãè äðàò àçí û é öåí ò ð
Í2Î
|
|
|
|
HS |
|
|
|
|
|
|
H |
H |
O |
|
Åí î è ë -ÀÏ Á |
|
|
|
|
(R)H 3C |
C |
S |
|
|
|
|
|
||
C |
C |
Í ÀÄÔÍ |
|
|
|
|
|||
|
|
|
|
|
2 |
|
|
|
|
|
|
|
|
ен о и л редукт азн ы й цен т р |
|
|
|
||
|
|
|
|
|
Í ÀÄÔ + |
|
H |
H |
O |
|
|
|
|
HS |
|
(R)H 3C |
C2 |
C2 |
C S |
|
H2 |
H2 |
O |
|
Àöè ë-ÀÏ Á |
|
|
Àöè ë-ÀÏ Á |
|
(R)H 3C |
C |
S |
|
|
|
|
HS |
||
C |
C |
Í 2Î |
|
|
|
||||
т и о эст еразн ы й цен т р |
|
|
|
|
|||||
HS |
ÀÏ Á |
|
|
|
|||||
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
HS |
|
|
|
|
RCÎÎÍ
ï àëüì è òàò
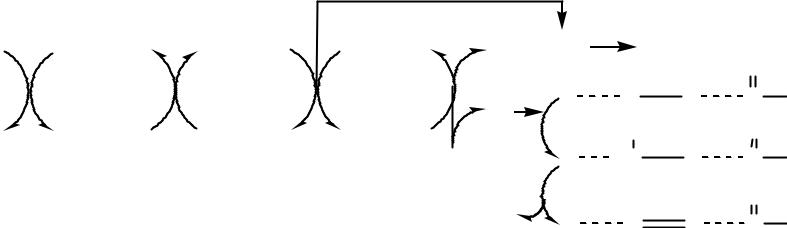
Десатуразы
Í ÀÄÍ 2 |
ÔÀÄ |
Fe3+ (b5) |
Fe3+ |
O2- + 2H+ |
H2O |
O |
|
||
|
Редуктаза |
Öèòî õðî ì |
Десатураза |
R |
H2 |
H2 |
C |
SKoA |
|
Í ÀÄ+ |
ÔÀÄÍ 2 |
Fe2+ (b5) |
Fe2+ |
O |
C |
C |
|||
|
OH |
H |
O |
|
|||||
|
|
|
|||||||
|
|
|
|
O2 |
R |
CH |
C2 |
C |
SKoA |
|
|
|
|
H2O |
|
H |
H |
O |
|
|
|
|
|
R |
C |
SKoA |
|||
|
|
|
|
C |
C |
||||
Элонгазы
1.Удлинение ЖК называется элонгацией.
2.Для каждой длины ЖК существуют свои элонгазы (16→ 18, 18 → 20, 20 → 22, 22 → 24). Последовательность реакций аналогична синтезу пальмитиновой кислоты, однако в данном случае синтез идет не на АПБ, а на КоА.
3.Основной продукт элонгации в печени — стеариновая кислота.
4.В нервных тканях образуются ЖК с длинной цепью (С=20-24), необходимые для синтеза сфинголипидов

Катаболизм ЖК
Ферментативный
•β-окисление (основной путь).
•α-окисление
•ω-окисление ЖК,
•деградация ЖК в пероксисомах
Неферментативный
Перекисное окисление липидов (ПОЛ)
•Хотя побочные пути (α-, ω-окисление ЖК, деградация ЖК в пероксисомах) количественно менее важны, их нарушение может приводить к тяжелым заболеваниям.
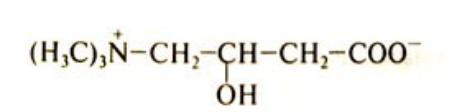
Транспорт ЖК в митохондрии
• ЖК с короткой и средней цепью (от 4 до 12 атомов С) могут проникать в матрикс митохондрий путём диффузии. Активация этих ЖК происходит ацил-КоА синтетазами в матриксе митохондрий.
• ЖК с длинной цепью, сначала активируются в цитозоле (ацил- КоА синтетазами на внешней мембране митохондрий), а затем переносятся в матрикс митохондрий специальной транспортной системой с помощью карнитина.
• Карнитин поступает с пищей или
синтезируется из лизина и |
|
|
метионина с участием витамина |
L-карнитин |
|
С. |
||
|
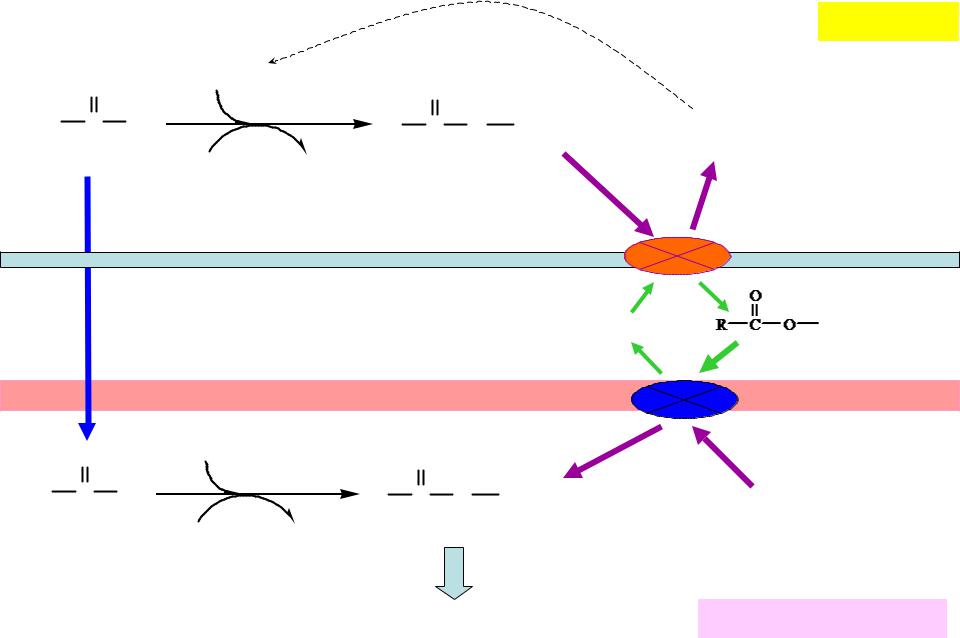
Транспорт ЖК в митохондрии
|
|
|
|
|
Цитоплазма |
O |
HS-KoA |
|
O |
|
|
|
|
|
|
||
R C OH |
|
|
R |
C O SKoA |
НS-КоА «-» Малонил-КоА |
Жирн ая кисло та |
ÀÒÔ |
ÀÌ Ô+ÔÔí |
Àöèë-SKoA |
||
|
|
|
Карнитинацил- |
||
|
àöèë-Êî À-ñèí ò åò àçà |
|
|||
|
|
|
|
|
трансфераза 1 |
|
|
|
|
Карнитин |
Карнитин |
|
HS-KoA |
|
Карнитинацил- |
|
O |
O |
трансфераза 2 |
||
|
||||
R C OH |
|
R C O SKoA |
НS-КоА |
|
Жирн ая кисло таÀÒÔ |
ÀÌ Ô+ÔÔí Àöèë-SKoA |
|||
|
||||
|
àöèë-Êî À-ñèí ò åò àçà |
|
||
|
|
β-окисление |
Матрикс митохондрии |
|
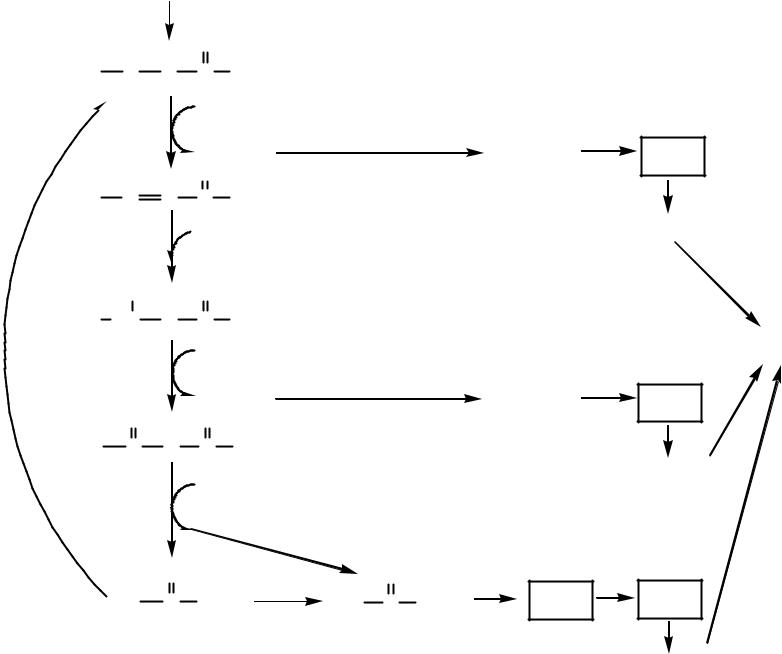
|
ï àëüì èòèë-Êî À C=16 |
Реакции β-окисление ЖК |
|||||
|
|
|
O |
|
|
||
R |
H |
H |
SKoA |
Àöèë-Êî À |
|
|
|
C2 |
C2 |
C |
|
|
|||
|
|
|
|
|
C=16,14,12,10,8,6,4 |
|
|
|
|
|
ÔÀÄ Àöèë-Êî À ÄÃ |
|
|
||
|
|
|
ÔÀÄÍ 2 |
|
7 ÔÀÄÍ 2 |
ÖÏ Ý |
|
|
H |
H |
O |
|
Åí î èë-Êî À |
|
|
R |
C |
SKoA |
|
|
|||
C |
C |
|
|
||||
|
|
|
Í 2Î |
Ен о ил-Ко А гидрат аза |
14 ÀÒÔ |
||
|
|
|
|
|
|||
|
ÎÍ |
H |
O |
|
|
|
|
|
R |
CÍ |
C2 |
C |
SKoA |
О ксиацил-Ко А |
|
||
7 цикло в |
|
|
Í ÀÄ+ |
О ксиацил-Ко А ДГ |
|
|||
|
|
|
|
|
||||
|
|
|
Í ÀÄÍ |
|
||||
|
Î |
|
2 |
|
|
7 Í ÀÄÍ 2 |
||
|
H2 |
O |
|
|
|
|
|
|
R |
C |
C |
SKoA |
Êåòî àöèë-Êî À |
|
|||
C |
|
|||||||
C=14,12,10, |
ÍSKoA |
|
|
|
|
|||
8,6,4 |
|
Êåò î àöèë-Êî À ò èî ëàçà |
|
|||||
|
|
|
|
|
|
|
||
|
|
O |
|
|
|
|
O |
ÖÒÊ |
|
R |
C |
SKoA |
C = 2 |
H3C C SKoA |
|||
|
|
Àöèë-Êî À |
8 Ацетил-Ко А |
|
||||
131 ÀÒÔ
ÖÏ Ý
21 ÀÒÔ
ÖÏ Ý
96 ÀÒÔ
β-окисление насыщенной ЖК с нечетным количеством атомов С
Последний цикл β-окисления сопровождается образованием не Ацетил-КоА, а Пропионил-КоА с 3 атомами С.
H2C CH3
CO
SKoA
Пропионил-КоА
|
|
|
|
|
|
COOH |
АТФАДФ+Фн |
COOH |
|
COOH |
CH2 |
||
|
|
HC CH3 |
H3C |
CH |
|
H2C |
|
биотин |
CO |
|
CO |
Â12 |
CO |
CO2 |
|
|
|
|||
SKoA |
|
SKoA |
SKoA |
|||
|
D-Метилмалонил-КоА |
L-Метилмалонил-КоА |
Сукцинил-КоА |
|||
Пропионил-КоА- |
Метилмалонил-КоА |
|
Метилмалонил-КоА |
|||
карбоксилаза |
-рацемаза |
|
|
-изомераза |
|
|
Метилмалоновая кислота
Повреждение
ЦНС
ÖÒÊ
ÖÏÝ
6 ÀÒÔ