

4.4. Особенности метаболизма в нервной ткани
De novo – метаболический принцип биохимии головного мозга. Кроме потребления основных питательных веществ и принципиально незаменимых для организма молекул, анаболизм мозга достаточно автономный. De novo синтез всех веществ в мозге начинается с глюкозы и аминокислот. Около 25% энергии тратится на анаболические процессы обновления белков и липидов.
Нервная ткань сама синтезирует белки, нуклеиновые кислоты и нуклеотиды. Синтез пуринов de novo самодостаточен, пиримидины предпочтительно транспортируются из крови.
Интенсивный аэробный катаболизм глюкозы происходит в основном за счет гликолиза, значительно меньше – пентозофосфатный путь. Цикл Кребса обеспечивает энергию для функционирования нервной ткани.
Обмен липидов является для мозга преимущественно внутренним и использует глюкозу в качестве предшественника; основным источником восстановительных эквивалентов является НАДФН-зависимая изоцитратдегидрогеназа. Для нервной ткани характерна высокая скорость обновления фосфолипидов. Обмен с липопротеинами крови в нервной ткани не осуществляется.
Катаболизм азота (цикл мочевины) в нервной ткани неполный, поэтому мозг чувствителен к повышению концентрации аммиака и выделяет азот в основном в форме глутамина (и других аминокислот).
Специфическими для нервной ткани являются метаболические пути синтеза и реутилизации ряда нейромедиаторов.
Обмен аммиака в головном мозге. В головном мозге глутаминсинтетаза, которая находится преимущественно в астроцитах, в значительной степени отвечает за удаление как полученного из крови, так и метаболически вырабатываемого аммиака.
Аммиак, поступающий в мозг путем диффузии из крови или спинномозговой жидкости, быстро фиксируется в астроцитах, преимущественно в форме глутамина. Фермент глутаминаза снова может выделять аммиак из глутамина. Невозможность выведения аммиака при его избытке в крови сдвигает метаболизм ЦНС в сторону исчерпания кетокислот, так как они используются для связывания аммиака; в свою очередь, это приводит к дефициту энергетического обмена, осмотическому дисбалансу, снижению скорости регенерации «метаболических» нейромедиаторов (ГАМК) и функциональному расстройству нервной системы.
Метаболизм аминокислот в головном мозге образует нейротрансмиттеры (нейромедиаторы), которые подвергаются вторичной переработке. Высокие концентрации аминокислот в нервных тканях и ликворе (намного выше, чем в крови, особенно для Глу, Гли, Глн, Асн, Сер) объясняются их интенсивным метаболическим оборотом как предшественников нейромедиаторов.
Астроциты превращают глутамат в глутамин и возвращают его обратно в нейрон для дальнейшего использования. Также, синтез глутамата и глутамина de novo интенсивно происходит в астроцитах головного мозга через метаболический путь пируваткарбоксилазы (этот фермент преимущественно локализован в астроцитах).
Особым метаболическим процессом нервной ткани является шунт гамма-аминомасляной кислоты. Шунт ГАМК – это замкнутый метаболический процесс, имеющий двойную цель – производство и сохранение запаса тормозного нейромедиатора ГАМК из глутаминовой кислоты - возбуждающего медиатора, который в свою очередь, может образовываться из ГАМК. ГАМК образуется декарбоксилированием глутаминовой кислоты (фермент декарбоксилаза глутаминовой кислоты). ГАМК-шунт – это процесс, характерный исключительно для нервной ткани, так как здесь ГАМК постоянно необходим в большом количестве как сигнальная молекула. Глюкоза является основным предшественником производства ГАМК in vivo, хотя пируват и другие аминокислоты также могут действовать как предшественники (через цикл Кребса). Реакции ГАМК-шунта (Рис. 27):
1. ГАМК образуется при декарбоксилировании глутамата с участием фермента глутаматдекарбоксилазы (кофермент пиридоксальфосфат).
2. Следующим шагом в шунте ГАМК является трансаминирование ГАМК и α-кетоглутарата (фермент ГАМК-α-кетоглутарат-трансаминаза, сокращенно ГАМК-Т) с образованием глутамата и янтарного полуальдегида:
α-кетоглутарат + ГАМК → L-глутамат+ янтарный полуальдегид
3. Затем янтарный полуальдегид восстанавливается до янтарной кислоты (сукцината) (фермент НАД-зависимая сукцинат-полуальдегиддегидрогеназа).
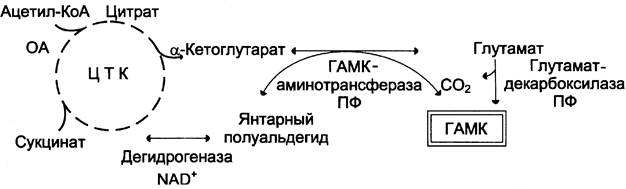
Рис.27. Реакции ГАМК-шунта [15].
4. Далее сукцинат через цикл Кребса превращается в α-кетоглутарат. Превращение α-кетоглутарата в глутамат – реакция восстановительного аминирования, катализируемая ферментом глутаматдегидрогеназой (кофермент НАДФН2).
В результате реакций данного процесса происходит как образование ГАМК (реакция 1), так и его распад (реакции 2 и 3). ГАМК-шунт связан с ЦТК метаболически: образованный при дегидрировании янтарного полуальдегида сукцинат может вовлекаться в цикл Кребса. Основное предназначение ГАМК-шунта – синтез и утилизация нейромедиатора ГАМК.
Витамин В6 в виде кофермента пиридоксальфосфата является незаменимым кофактором при синтезе ГАМК в тканях мозга, а также и переаминирования ГАМК с альфа-кетоглутаратом при образовании возбуждающего медиатора – глутаминовой кислоты, что объясняет нарушения работы нервной системы при гиповитаминозах пиридоксина.